
Dinosaures
| Dinosauria | |
|---|---|
 D'esquerra a dreta i de dalt a baix: dos plateosaures; un estiracosaure i un escolosaure; un estruç; un barosaure i dos al·losaures; diversos hadrosàurids; i Sinocalliopteryx gigas | |
| Dades | |
| Font de | petjada de dinosaure |
| Període | |
| Taxonomia | |
| Super-regne | Holozoa |
| Regne | Animalia |
| Fílum | Chordata |
| Ordre | Eotetrapodiformes |
| Classe | Reptilia |
| Superordre | Dinosauria Owen, 1842 |
| Nomenclatura | |
| Significat | Llangardaixos terribles |
| Ordres i subordres | |
Els dinosaures (Dinosauria) són un grup heterogeni de sauròpsids. Se sap que aparegueren durant el període Triàsic, fa entre 233 i 243 milions d'anys, tot i que se'n continuen estudiant els orígens i l'evolució. Després de l'extinció del Triàsic-Juràssic, fa aproximadament 201,3 milions d'anys, es convertiren en els vertebrats terrestres dominants, condició que mantingueren durant el Juràssic i el Cretaci. El registre fòssil demostra que els ocells són dinosaures amb plomes, que evolucionaren a partir de teròpodes més primitius durant el Juràssic superior i que són l'únic grup de dinosaures que no desaparegué en l'extinció del Cretaci-Paleogen, fa aproximadament 66 milions d'anys. Així doncs, els dinosaures es poden dividir en «aviaris» (els ocells) i «no aviaris» (tota la resta).
Exhibeixen una gran variabilitat taxonòmica, morfològica i ecològica. Hi ha més de 10.000 espècies vivents d'ocells, cosa que els situa entre els grups de vertebrats més diversos del món. Els paleontòlegs han descrit més de 900 gèneres i 1.000 espècies de dinosaures no aviaris a partir de fòssils. Hi ha dinosaures vivents (ocells) i fòssils als set continents. Fins ben entrat el segle xx, quan els ocells foren reclassificats com a dinosaures, la major part de la comunitat científica veia aquests últims com a animals lents i de sang freda. Tanmateix, el consens que ha emergit dels estudis duts a terme des de la dècada del 1970 és que eren criatures actives, amb un ritme metabòlic alt i nombroses adaptacions per a la interacció social. N'hi havia d'herbívors i de carnívors. Les proves fan pensar que tots els dinosaures ponen ous i que molts d'ells, tant aviaris com no aviaris, fan nius.
El descobriment el 1860 de l'arqueòpterix fou el primer a suggerir una relació estreta entre els dinosaures i els ocells; a part de la presència d'empremtes fossilitzades de plomes, l'arqueòpterix era molt similar al petit dinosaure carnívor contemporani Compsognathus. La recerca duta a terme des de la dècada del 1970 indica que els dinosaures teròpodes són els avantpassats més versemblants dels ocells; de fet, la majoria de científics veuen els ocells com a únics dinosaures supervivents, i alguns, fins i tot, creuen que els dinosaures i els ocells haurien de ser agrupats en una única classe biològica.[2] Els crocodilians són els altres parents propers dels dinosaures supervivents, i ambdós grups són membres del clade dels arcosaures, un grup de rèptils que aparegué per primer cop al Permià molt tardà i que esdevingué dominant a mitjans del Triàsic.
Els dinosaures no sempre visqueren en climes càlids com s'ha cregut tradicionalment. A partir del segle xxi es trobaren fòssils de dinosaures del principi del Cretaci (entre 125 i 110 milions d'anys enrere) al nord de la Xina que demostren que també vivien sota hiverns molt freds; això explicaria la presència d'una mena de plomes (estructures filamentoses de diversos tipus) en certs tipus de dinosaures d'aquella zona i època.[3]
Des que a principi del segle xix es reconegueren els primers fòssils de dinosaure, els esquelets de dinosaure muntats han esdevingut importants atraccions a museus d'arreu del món. Els dinosaures han esdevingut part de la cultura global i resten populars entre els nens i els adults. Han aparegut en llibres i pel·lícules molt vistes (notablement Parc Juràssic), i els nous descobriments són habitualment coberts pels mitjans de comunicació.
El terme «dinosaure» fou encunyat originalment el 1842 per Richard Owen i prové dels mots grecs δεινός (dinós), ‘terrible’, ‘poderós’, ‘imponent’, i σαῦρος (sauros), ‘llangardaix’. A vegades s'usa el terme de manera informal per descriure altres rèptils prehistòrics, com ara el pelicosaure Dimetrodon, els pterosaures alats, i els ictiosaures, plesiosaures i mosasàurids aquàtics, tot i que cap d'aquests grups no eren dinosaures.
Morfologia

Els dinosaures (a part dels ocells) poden ser descrits com a rèptils arcosaures terrestres amb les potes mantingudes erectes sota el cos, que existiren des del Triàsic superior (apareixent per primer cop a l'estatge faunístic Carnià) fins al Cretaci superior (extingint-se a finals del Maastrichtià).[4] Molts animals prehistòrics són popularment considerats dinosaures, com ara els ictiosaures, mosasaures, plesiosaures, pterosaures o Dimetrodon, però no se'ls classifica científicament com a tals. Els rèptils marins com els ictiosaures, mosasaures i plesiosaures no eren ni terrestres ni arcosaures; els pterosaures eren arcosaures però no terrestres i el dimetrodont era un animal del Permià més proper als mamífers.[5] Els dinosaures foren els vertebrats terrestres dominants del Mesozoic, especialment el Juràssic i el Cretaci. Els altres grups d'animals quedaren limitats en mida i en nínxols; els mamífers, per exemple, rarament sobrepassaven la mida d'un gat, i eren generalment carnívors de la mida de rosegadors que caçaven preses petites.[6] Una excepció notable és Repenomamus giganticus, un triconodont que pesava entre 12 i 14 kg i que se sap que es menjava petits dinosaures com ara cries de psitacosaure.[7]
Els dinosaures eren un grup d'animals extremament variat; segons un estudi del 2006, fins al present s'han identificat amb certesa més de 500 gèneres de dinosaures, i el nombre total de gèneres preservats en el registre fòssil ha estat estimat en, aproximadament, 1.850, gairebé tres quarts dels quals encara han de ser descoberts.[8] Un estudi anterior estimà que existiren aproximadament 3.400 gèneres de dinosaures, incloent-n'hi molts que no haurien quedat preservats en el registre fòssil.[9] Alguns eren herbívors i altres eren carnívors. Alguns dinosaures eren bípedes, altres quadrúpedes, i altres, com l'ammosaure o Iguanodon, podien caminar indistintament a dues o a quatre potes. Molts tenien una armadura òssia, o modificacions cranials com ara banyes i crestes. Tot i que se'ls coneix per la gran mida, en realitat la majoria de dinosaures eren de mida humana o més petits. Se n'han trobat restes a tots els continents de la Terra, incloent-hi l'Antàrtida.[10] No es coneixen dinosaures mesozoics que hagin viscut en hàbitats marins o aeris, tot i que és possible que alguns teròpodes amb plomes poguessin volar.
Trets distintius
Tot i que els descobriments recents han fet que sigui més difícil presentar una llista dels trets distintius dels dinosaures amb consens universal, gairebé tots els dinosaures descoberts fins ara comparteixen certes modificacions de l'esquelet ancestral dels arcosaures. Tot i que alguns grups posteriors de dinosaures presentaven versions encara més modificades d'aquests trets, són considerats típics de tots els dinosaures; els dinosaures més primitius els tenien i els passaren a tots els seus descendents. Aquestes característiques comunes d'un grup taxonòmic reben el nom de sinapomorfies.
Les sinapomorfies dels dinosaures inclouen una cresta allargada a l'húmer, per acomodar l'ancoratge dels músculs deltopectorals; una base a la part posterior de l'ili; una tíbia amb una vora inferior ampla i una brida que apunta cap endarrere i cap enfora i una projecció ascendent de l'astràgal, un dels ossos del turmell, que l'encaixa amb la tíbia.[11]

Molts dinosaures compartien una varietat d'altres característiques esqueletals. Tanmateix, com que també eren comunes en altres grups d'arcosaures, o no estaven presents en tots els dinosaures primerencs, aquestes característiques no són considerades sinapomorfies. Per exemple, sent rèptils diàpsids, els dinosaures tenien dos parells de finestres temporals (obertures cranials darrere els ulls), i com a membres del grup de diàpsids dels arcosaures, tenien obertures addicionals al musell i maxil·lar inferior.[12] A més, actualment se sap que algunes característiques, antigament considerades sinapomorfies, en realitat aparegueren abans dels dinosaures, o eren absents en els dinosaures més primitius i evolucionaren independentment en diferents grups de dinosaures. Aquests trets inclouen un omòplat allargat; un sacre compost de tres o més vèrtebres fusionades (altres arcosaures també en tenen tres, però l'herrerasaure en tenia únicament dues);[11] i un acetàbul amb un forat al centre de la seva superfície interior (que queda tancat en alguns dinosaures com ara Saturnalia).[13] Una altra dificultat a l'hora de determinar característiques distintives dels dinosaures és que els dinosaures primitius i altres arcosaures del Triàsic superior sovint són poc coneguts i s'assemblen en moltes maneres; a vegades s'han identificat erròniament aquests animals en la literatura científica.[14]

Els dinosaures es movien erectes de manera similar a la majoria de mamífers moderns, però diferent a la majoria d'altres rèptils, que tenen les potes plegades a cada banda.[15] La seva postura es devia al desenvolupament d'un recés enfocat vers el lateral a la pelvis (normalment una ranura oberta), i el corresponent cap del fèmur enfocat vers l'interior.[16] La seva postura erecta permetia als dinosaures respirar amb facilitat mentre es movien, permetent nivells de resistència i d'activitat físiques que superaven els dels rèptils amb «potes plegades».[17] Les potes erectes probablement també contribuïren a l'evolució de dinosaures de grans dimensions reduint l'estrès de flexió a les potes.[18] Alguns arcosaures no dinosaures, com ara els rauisucs, també tenien membres erectes, però ho aconseguien per mitjà d'una configuració de «pilar erecte» de l'articulació del maluc, en què en lloc de tenir una projecció del fèmur que s'inseria en una ranura del maluc, l'os ili estava girat per formar una base superjacent.[18]
Taxonomia
El tàxon Dinosauria fou descrit formalment el 1842 pel paleontòleg anglès Richard Owen, que l'utilitzà per denominar la "distinta tribu o subordre dels rèptils saurians", que aleshores estaven sent reconeguts a Anglaterra i arreu del món.[19] El terme prové dels mots grecs δεινός (deinos) "terrible, poderós, imponent" i σαῦρος (sauros) "llangardaix" o "rèptil".[20] Tot i que el nom taxonòmic ha estat sovint interpretat com una referència a les dents, urpes i altres característiques temibles dels dinosaures, Owen simplement volia evocar la seva mida i majestuositat.[21]

En la taxonomia filogenètica, els dinosaures són definits generalment com a tots els descendents de l'avantpassat comú més recent de Triceratops i els ocells moderns.[22] També s'ha suggerit que Dinosauria sigui definit com tots els descendents de l'avantpassat comú més recent del megalosaure i d'Iguanodon, que foren dos dels tres gèneres citats per Richard Owen quan descrigué els dinosaures.[23] Ambdues definicions resulten en el reconeixement com a dinosaures del mateix conjunt d'animals, incloent-hi els teròpodes (majoritàriament carnívors bípedes), sauropodomorfs (majoritàriament grans quadrúpedes herbívors), estegosaures (herbívors quadrúpedes amb plaques), ceratops (quadrúpedes herbívors amb banyes i crestes), i ornitòpodes (herbívors bípedes o quadrúpedes, incloent-hi els hadrosaures). Aquestes definicions estan escrites per correspondre's amb els conceptes científics de dinosaures anteriors a l'ús modern de la filogènia. Aquesta continuïtat de significat intenta evitar la confusió quant al que vol dir el terme "dinosaure".
Hi ha un consens gairebé universal entre els paleontòlegs que els ocells són descendents de dinosaures teròpodes. Utilitzant l'estricta definició cladística que exigeix que tots els descendents d'un mateix avantpassat comú siguin inclosos en un grup per tal que aquest sigui natural, els ocells són dinosaures, i, per tant, els dinosaures no estan extints. Els ocells són classificats per la majoria de paleontòlegs com a membres del subgrup dels maniraptors, que són celurosaures, que són teròpodes, que són saurisquis, que són dinosaures.[24]
Des del punt de vista cladístic, els ocells són dinosaures, però en la parla vulgar, el terme «dinosaure» no els inclou. A més, referir-se als ocells que no són dinosaures com a "dinosaures no aviaris" és maldestre. Per qüestions de claredat, aquest article utilitza "dinosaure" com a sinònim de "dinosaure no aviari". El terme "dinosaure no aviari" serà utilitzat per emfatitzar el que sigui necessari. També és tècnicament correcte referir-se als dinosaures com un grup distint en el sistema de classificació linneana, més antic, que accepta tàxons parafilètics que excloguin alguns descendents d'un avantpassat comú.
Història natural
Orígens i primera fase d'evolució

Durant molt de temps, molts científics pensaren que els dinosaures eren polifilètics, amb múltiples grups de "dinosaures" no relacionats que havien evolucionat en resposta a pressions similars,[25][26][27] però actualment se sap que els dinosaures formen un únic grup.[28][11]
Els dinosaures divergiren dels seus avantpassats arcosaures fa aproximadament 230 milions d'anys, vers el Triàsic mitjà-superior, aproximadament vint milions d'anys després que l'extinció permiana hagués exterminat al voltant del 95% de totes les formes de vida de la Terra.[29][30] La datació radiomètrica de la formació rocosa que contenia els fòssils del dinosaure primerenc Eoraptor, demostra la seva presència en el registre fòssil en aquell temps. Els paleontòlegs creuen que Eoraptor és similar a l'avantpassat comú de tots els dinosaures;[31] si això és cert, les seves característiques suggereixen que els primers dinosaures eren petits depredadors bípedes.[32] El descobriment d'ornitodirs primitius semblants a dinosaures com ara Marasuchus o Lagerpeton en estrats del Triàsic mitjà de l'Argentina dona suport a aquesta visió: anàlisis de fòssils descoberts suggereixen que aquests animals eren en efecte petits depredadors bípedes.
Quan aparegueren els dinosaures, els hàbitats terrestres estaven ocupats per diversos tipus d'arcosaures basals i teràpsids, com ara etosaures, cinodonts, dicinodonts, ornitosúquids, rauisucs i rincosaures. La majoria d'aquests animals s'extingiren durant el Triàsic, en un dels dos esdeveniments següents:
- El primer, aproximadament al límit entre els estatges faunístics Carnià i Norià (fa 215 milions d'anys), s'extingiren els dicinodonts i una varietat d'arcosauromorfs basals, incloent-hi els prolacertiformes i els rincosaures.
- El segon esdeveniment fou l'extinció massiva del Triàsic-Juràssic (fa uns 200 milions d'anys), que significà la fi de la majoria d'altres grups d'arcosaures primitius, com ara els etosaures, ornitosúquids, fitosaures i rauisucs.
Aquestes desaparicions deixaren una fauna terrestre composta de crocodilomorfs, dinosaures, mamífers, pterosaures i tortugues.[11]
Els primers llinatges de dinosaures primitius es diversificaren durant els estatges faunístics del Triàsic Carnià i Norià, probablement ocupant els nínxols de grups que s'extingien. Tradicionalment, es pensava que els dinosaures havien substituït altres animals terrestres del Triàsic imposant-se al final d'un llarg període de competició. Actualment, això sembla inversemblant per una sèrie de raons. Els dinosaures no presenten un patró d'augment constant en diversitat ni en nombre, com seria d'esperar si haguessin substituït per mitjà de la competició altres grups; tot i així, eren molt rars durant el Carnià, representant únicament un 1-2% dels animals presents en la fauna. En el Norià, en canvi, després de l'extinció de diversos altres grups, esdevingueren una part significativa de la fauna, representant-ne un 50-90% dels individus. A més, actualment se sap que el que havia estat considerat una adaptació clau dels dinosaures, la seva postura erecta, també estava present en diversos grups de la mateixa època que no tingueren èxit, com etosaures, ornitosúquids, rauisucs, i alguns grups de crocodilmorfs. Finalment, el Triàsic superior fou una època de gran inestabilitat en la vida a la Terra, amb canvis en la vida vegetal, la vida marina i el clima.[11]
Baixa diversificació durant el Cretaci
Anàlisis estadístiques basades en dades brutes suggereixen que els dinosaures es diversificaren al Cretaci superior; és a dir, que n'augmentà el nombre d'espècies. Tanmateix, al juliol de 2008, Graeme T. Lloyd et al. argumentaren que aquesta diversificació aparent era una il·lusió causada per un biaix de mostres, car les roques del Cretaci superior han estat estudiades molt intensament. En canvi, segons ells, els dinosaures només experimentaren dues diversificacions significatives al Cretaci superior, les radiacions inicials dels euhadrosaures i els ceratops. Al Cretaci mitjà, les plantes angiospermes amb flor esdevingueren un element clau dels ecosistemes terrestres, que anteriorment havien estat dominats per gimnospermes, com ara coníferes. Els copròlits o excrements fossilitzats dels dinosaures indiquen que, mentre que alguns d'ells menjaven angiospermes, la majoria de dinosaures herbívors s'alimentaven principalment de gimnospermes. Mentrestant, els insectes i mamífers herbívors es diversificaren ràpidament per aprofitar el nou tipus d'aliment vegetal, alhora que els llangardaixos, serps, crocodilians i ocells també es diversificaven. Lloyd et al. suggereixen que el fracàs dels dinosaures a l'hora de diversificar-se mentre els ecosistemes canviaven els condemnà a l'extinció.[33]
Classificació
Els dinosaures (incloent-hi les aus) són arcosaures, com els crocodilians moderns. Els cranis diàpsids dels arcosaures tenen dos forats, denominats finestres temporals, situades on s'ancoren els músculs mandibulars, i una finestra antorbitària addicional davant dels ulls. La majoria de rèptils, incloent-hi les aus, són diàpsids, mentre que els mamífers, amb una única finestra temporal, reben el nom de sinàpsids. Anatòmicament, els arcosaures presenten moltes altres característiques dels arcosaures, incloent-hi dents que creixen de ranures en lloc de ser extensions directes dels ossos maxil·lars. Dins del grup dels arcosaures, els dinosaures es distingeixen sobretot per la seva marxa. Les potes dels dinosaures s'alcen directament sota el cos, mentre que les dels llangardaixos i cocodrils es pleguen cap als costats.
Col·lectivament, els dinosaures són habitualment considerats un superordre o un clade sense rang. Se'ls subdivideix en dos ordres, els saurisquis i els ornitisquis], segons l'estructura de la pelvis. Els saurisquis inclouen els tàxons que comparteixen un avantpassat comú més recent amb els ocells que amb els ornitisquis, mentre que els ornitisquis inclouen tots els tàxons que comparteixen un avantpassat comú més recent amb Triceratops que amb els saurisquis. Els saurisquis ('maluc de llangardaix', del grec sauros (σαυρος) 'llangardaix' i iskhion (ισχιον) 'articulació del maluc') retingueren l'estructura pelviana dels seus avantpassats, amb un pubis apuntant cranialment (vers el crani) o cap endavant.[16] Aquesta forma bàsica fou modificada en alguns grups (l'herrerasaure,[34] els terezinosauroïdeus,[35] els dromeosàurids,[36] i els ocells[24]) rotant el pubis cap enrere en certa manera. Els saurisquis inclouen els teròpodes (bípedes i en gran part carnívors, tret dels ocells) i els sauropodomorfs (herbívors quadrúpedes de coll llarg).
En canvi, els ornitisquis (‘maluc d'ocell’, del grec ornithios (ορνιθειος) ‘aviari’ i iskhion (ισχιον) ‘maluc’) tenien una pelvis que s'assemblava superficialment a la dels ocells: el pubis estava orientat caudalment, és a dir, cap enrere. A diferència dels ocells, el pubis dels ornitisquis també solia tenir un procés addicional orientat cap endavant; els ornitisquis incloïen una varietat d'herbívors. Tanmateix, i malgrat el nom dels grups, cal recordar que els ocells evolucionaren de dinosaures saurisquis, és a dir, amb "maluc de llangardaix".
-
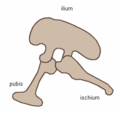 Estructura pelviana dels saurisquis (costat esquerre)
Estructura pelviana dels saurisquis (costat esquerre) -
 Pelvis de tiranosaure (estructura pelviana de saurisqui - costat esquerre)
Pelvis de tiranosaure (estructura pelviana de saurisqui - costat esquerre) -
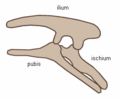 Estructura pelviana dels ornitisquis (costat esquerre).
Estructura pelviana dels ornitisquis (costat esquerre). -
 Pelvis d'edmontosaure (estructura pelviana d'ornitisqui - costat esquerre)
Pelvis d'edmontosaure (estructura pelviana d'ornitisqui - costat esquerre)




Aquesta és una classificació simplificada de les famílies de dinosaures:

- Dinosauria:
- Saurischia (teròpodes i sauròpodes).
- Herrerasauria (predadors bípedes primitius).
- Theropoda (tots bípedes; la majoria eren carnívors).
- Coelophysoidea (Coelophysis i parents propers).
- Ceratosauria (Ceratosaurus i abelisàurids - aquests últims foren importants depredadors durant el Cretaci superior als continents meridionals).
- Spinosauroidea (cossos llargs, braços curts, alguns d'ells amb cranis "crocodilians" i "veles" òssies a l'esquena).
- Carnosauria (Allosaurus i parents propers, com ara Carcharodontosaurus).
- Coelurosauria (diversos, amb una varietat de mida i de nínxols).
- Tyrannosauroidea (gran varietat de mida, sovint amb potes davanteres reduïdes).
- Ornithomimosauria ("imitadors d'estruços", sense dents, carnívors i herbívors).
- Therizinosauroidea (herbívors bípedes amb braços llargs i un cap petit).
- Alvarezsauria (Alvarezsaurus i parent propers).
- Oviraptorosauria (sense dents; de dieta i estil de vida incerts).
- Dromaeosauridae (com els "raptors" de Parc Juràssic).
- Troodontidae (similars als dromeosàurids, però amb una fisonomia més lleugera, i possiblement omnívors).
- Avialae (els únics dinosaures vivents).

- Sauropodomorpha (herbívors quadrúpedes amb cap petit i coll i cua llargs, amb cossos semblants al dels elefants).
- "Prosauropoda" (parents primitius dels sauròpodes; de mida petita a mida bastant gran; algun d'ells possiblement omnívors, bípedes i quadrúpedes).
- Sauropoda (molt grans, habitualment de més de 15 metres de llarg).
- Diplodocoidea (crani i cua allargats; dents típicament estretes i semblants a llapis).
- Macronaria (crani amb forma de caixa; dents amb forma de cullera o de llapis).
- Brachiosauridae (colls molt llargs; potes anteriors més curtes que les posteriors).
- Titanosauria (diversos; pesants, amb malucs amples; especialment comuns als continents meridionals durant el Cretaci superior).

- Ornithischia (diversos herbívors quadrúpedes i bípedes).
- Heterodontosauridae (herbívors i omnívors de mida aproximada d'un metre amb dents canines prominents).
- Thyreophora (dinosaures blindats, majoritàriament quadrúpedes).
- Ankylosauria (blindats amb escuts; alguns tenien cues semblants a maces).
- Stegosauria (blindats amb punxes i plaques).
- Ornithopoda (diversos, des d'un metre de mida fins a 12 metres; animals que podien moure's de manera bípeda o quadrúpeda, amb un mètode masticatori que aprofitava la flexibilitat del crani i el gran nombre de dents)
- Hadrosauridae ("dinosaures de bec d'ànec").
- Pachycephalosauria ("caps d'os", bípedes amb una cúpula òssia al cap).
- Ceratopsia (dinosaures amb banyes i crestes, tot i que la majoria de formes primitives només tenien inicis d'aquestes característiques).
El 2017, un trio de científics britànics plantejaren una nova hipòtesi que reparteix els dinosaures en els clades Ornithoscelida (Ornithischia + Theropoda) i Saurischia (redefinit com a Sauropodomorpha + Herrerasauridae). Aquesta nova classificació, potencialment revolucionària, encara ha de ser avaluada per la resta de la comunitat científica.[37]
Evolució i paleobiogeografia
L'evolució dels dinosaures després del Triàsic segueix els canvis en la vegetació i en la situació dels continents. Al Triàsic superior i Juràssic inferior, els continents estaven units en una única massa terrestre anomenada Pangea, i hi havia una fauna global de dinosaures composta principalment de carnívors celofisoïdeus i herbívors prosauròpodes.[38] Les plantes gimnospermes (particularment les coníferes), una font d'aliment potencial, radiaren a finals del Triàsic. Els prosauròpodes no tenien mecanismes sofisticats per processar l'aliment a la boca, de manera que devien utilitzar altres mitjans de fragmentar el menjar en un punt posterior del sistema digestiu.[39] L'homogeneïtat general de les faunes de dinosaures continuà durant el Juràssic mitjà i superior, on la majoria d'indrets tenien depredadors consistents en ceratosaures, espinosauroïdeus i carnosaures, i herbívors consistents en ornitisquis estegosaurians i grans sauròpodes. En són exemples la formació de Morrison de Nord-amèrica i els estrats de Tendaguru de Tanzània. Els dinosaures xinesos presentaven algunes diferències, amb teròpodes sinraptòrids especialitzats i sauròpodes de coll llarg inusuals com ara el mamentxisaure.[38] Els ancilosaures i els ornitòpodes també esdevingueren més comuns, però els prosauròpodes s'extingiren. Els conífers i els pteridofitins eren les plantes més comunes. Els sauròpodes, igual que els prosauròpodes, no processaven l'aliment a la boca, però els ornitisquis començaren a desenvolupar diversos mètodes de processar el menjar a la boca, incloent-hi potencials òrgans semblants a galtes per mantenir l'aliment al menjar, i mocions de la mandíbula per moldre l'aliment.[39] Un altre esdeveniment evolutiu de gran importància del Juràssic fou l'aparició dels ocells autèntics, descendents de celurosaures maniraptors.[24]

A principis del Cretaci, amb la fragmentació de Pangea, els dinosaures començaren a diferir significativament d'una massa terrestre a l'altra. La primera part d'aquest període veié l'expansió dels ancilosaures, iguanodonts i braquiosàurids a Europa, Nord-amèrica i el nord d'Àfrica. Més endavant, a Àfrica, aquests dinosaures foren complementats o substituïts per grans teròpodes espinosàurids i carcarodontosàurids, i per sauròpodes rebbachisàurids i titanosaures, que també vivien a Sud-amèrica. A Àsia, celurosaures maniraptors com ara els dromeosàurids, troodòntids i oviraptorosaures esdevingueren els teròpodes més comuns, i els ancilosàurids i ceratops primitius com el psitacosaure esdevingueren herbívors importants. Mentrestant, Austràlia albergava una fauna d'ancilosaures basals, hipsilofodonts i iguanodonts.[38] Sembla que els estegosaures s'extingiren en algun moment a finals del Cretaci inferior o principis del Cretaci superior. Un canvi significatiu del Cretaci inferior, que seria amplificat al Cretaci superior, fou l'evolució de plantes amb flor. Al mateix temps, diversos grups de dinosaures herbívors desenvoluparen maneres més sofisticades de processar l'aliment a la boca: els ceratops desenvoluparen un mètode de tallar amb les dents apilades les unes sobre les altres en bateries, i els iguanodonts refinaren un mètode de moldre el menjar amb bateries de dents, portat a l'extrem en els hadrosàurids;[39] alguns sauròpodes també desenvoluparen bateries de dents, exemplificades clarament pel rebbachisàurid Nigersaurus.[40]
Al Cretaci superior hi havia tres faunes de dinosaures generals. Als continents septentrionals de Nord-amèrica i Euràsia els teròpodes més importants eren els tiranosàurids i diversos tipus de teròpodes maniraptors més petits, amb un conjunt herbívor predominantment ornitisqui amb hadrosàurids, ceratopsis, anquilosùrids i paquicefalosaures. Als continents meridionals, que formaven el supercontinent en procés de fragmentació Gondwana, els abelisàurids eren els teròpodes més comuns, i els sauròpodes titanosaurians els herbívors més habituals. Finalment, a Europa, els dromeosàurids, iguanodonts rabdodòntids, ancilosaures nodosàurids, i sauròpodes titanosaurians eren prevalents.[38] Les plantes amb flor estaven tenint una gran radiació,[39] amb l'aparició de les primeres poàcies a finals del Cretaci.[41] Els hadrosàurids amb dents moledores i els ceratops amb dents talladores es diversificaren molt a Nord-amèrica i Euràsia. Els teròpodes també passaven una radiació com a herbívors o omnívors, esdevenint comuns els terizinosaures i ornitomimosaures.[39]
L'extinció del Cretaci-Paleogen, que tingué lloc, fa aproximadament, 65 milions d'anys a finals del període Cretaci, causà l'extinció de tots els dinosaures tret dels ocells. Alguns altres grups de diàpsids, com ara els crocodilians, llangardaixos, serps, esfenodonts i coristoders, també sobrevisqueren a aquesta extinció (tot i que aquests últims s'extingirien a principis del Miocè).[42]
Paleobiologia
El coneixement sobre els dinosaures prové d'una varietat de registres fòssils i no fòssils, incloent-hi ossos fossilitzats, fems fossilitzats, petges, gastròlits, plomes, impressions de pell, òrgans interns, i teixits tous.[43][44] Molts camps de la ciència contribueixen al coneixement sobre els dinosaures, incloent-hi la física (especialment la biomecànica), la química, la biologia, i les ciències de la Terra (de les quals la paleontologia és una subdisciplina). Dos temes de particular interès i estudi han estat la mida i el comportament dels dinosaures.
Mida

Tot i que els indicis no són complets, queda clar que els dinosaures, com a grup, eren grans. Fins i tot per ser dinosaures, els sauròpodes eren enormes. Durant gran part de l'era dels dinosaures, els sauròpodes més petits eren més grossos que qualsevol altra cosa amb què compartien l'hàbitat, i els més grossos eren un ordre de magnitud més grossos que qualsevol altre animal que hi hagi hagut a la Terra des d'aleshores. Mamífers prehistòrics gegants com ara el paracerateri o el mamut colombí haurien semblat petits al costat dels enormes sauròpodes, i només uns quants animals aquàtics moderns s'acosten a la seva mida o la sobrepassen – l'exemple més notable és el rorqual blau, que arriba fins a un pes de 173 tones i una longitud de 30 metres.[45] S'han proposat diversos avantatges de la gran mida dels sauròpodes, incloent-hi protecció contra els depredadors, reducció de l'ús d'energia, i longevitat, però podria ser que l'avantatge més important fos en la dieta. Els animals grans tenen una digestió més eficient que la dels petits, car l'aliment roman més temps en el seu aparell digestiu. Això també els permet sobreviure amb aliments de valor nutritiu inferior que els animals més petits. Les restes de sauròpodes se solen trobar en formacions rocoses interpretades com a seques o estacionalment seques, i la capacitat de menjar grans quantitats de vegetació poc nutritiva hauria representat un avantatge en aquests ambients.[46]
Tanmateix, la majoria de dinosaures eren molt més petits que els grans sauròpodes. Els indicis actuals suggereixen que la mida mitjana dels dinosaures varià durant els períodes Triàsic, Juràssic inferior, Juràssic superior i Cretaci.[31] Els dinosaures teròpodes, quan se'ls classifica segons el pes estimat en categories basades en l'ordre de magnitud, sovint cauen en la categoria de 100 a 1.000 kg, mentre que els depredadors carnívors recents es troben en la categoria de 10 a 100 kg.[47] La moda de les masses corporals dels dinosaures és d'entre una i deu tones mètriques.[48] Això contrasta marcadament amb la mida dels mamífers del Cenozoic, que el Museu Nacional d'Història Natural estima entre dos i cinc quilograms.[49]
Extrems de mida
Només un percentatge minúscul dels animals arriba a fossilitzar-se, i la majoria d'ells romanen enterrats. Pocs dels exemplars que es descobreixen són esquelets complets, i les impressions de pell i altres teixits tous són rares. Reconstruir un esquelet complet comparant-ne la mida i la morfologia dels ossos a les d'espècies similars més ben conegudes és un art inexacte, i la reconstrucció dels músculs i altres òrgans de l'animal en vida és, en el millor dels casos, un joc de suposicions. Com a resultat d'això, els científics probablement no estaran mai segurs de quins foren els dinosaures més grossos i més petits.

El dinosaure més alt i més pesant conegut a partir d'esquelets bons és el braquiosaure (també conegut com a Giraffatitan). Se'n descobriren les restes a Tanzània entre el 1907 i el 1912. Els ossos de múltiples exemplars de dimensions similars foren incorporats en l'esquelet actualment muntat i exposat al Museu Humboldt de Berlín;[50] aquest esquelet mesura 12 metres d'alt i 22,5 de llargada, i pertanyia a un animal que pesava entre trenta i seixanta tones. El dinosaure complet més llarg és un Diplodocus, de 27 metres de llarg que fou descobert a Wyoming (Estats Units) i que fou exposat al Carnegie Natural History Museum de Pittsburgh el 1907.
Hi hagué dinosaures més grossos, però el coneixement sobre ells es basa únicament en un petit nombre de fòssils fragmentaris. La majoria dels exemplars d'herbívors més grossos foren descoberts a la dècada del 1970 o més tard, i inclouen el massiu argentinosaure, que podria haver pesat entre 80 i 100 tones; alguns dels més llargs, Diplodocus hallorum (antigament Seismosaurus) de 33,5 metres de llarg, i el supersaure, de 33 metres de llarg;[51] i el més alt, Sauroposeidon, que amb els seus 18 metres d'alçada podria haver arribat a la finestra d'un cinquè pis. El més llarg de tots podria haver estat Maraapunisaurus fragillimus, conegut únicament a partir d'un arc neural vertebral parcial actualment perdut, descrit el 1878. Extrapolant a partir de la il·lustració d'aquest os, l'animal podria haver mesurat 58 metres de llarg i haver pesat més de 120 tones,[46] sent més pesant que qualsevol altre dinosaure conegut, exceptuant, potser, el poc conegut Bruhathkayosaurus, que podria haver pesat entre 175 i 220 tones. El dinosaure carnívor més gran conegut fou l'espinosaure, que assolia mides d'entre 16 i 18 metres i un pes de 8.150 kg.[52] Altres carnívors grans inclouen el giganotosaure, el mapusaure, el tiranosaure i el carcarodontosaure.
Deixant de banda els ocells moderns, els dinosaures més petits coneguts tenien la mida aproximada d'un corb o d'una gallina. Els teròpodes Microraptor i Parvicursor mesuraven ambdós menys de 60 cm de llarg.
Comportament

Les interpretacions del comportament dels dinosaures es basen, generalment, en la postura dels fòssils i en l'hàbitat, en simulacions per ordinador de la seva biomecànica, i en comparacions amb animals actuals que visquin en nínxols ecològics similars. Per tant, el saber actual sobre el comportament dels dinosaures es basa en l'especulació, i probablement continuarà sent controvertit en el futur previsible. Tanmateix, existeix un consens general que alguns comportaments que són habituals entre els cocodrils i els ocells (els parents dels dinosaures més propers encara vivents) també eren habituals entre els dinosaures.
El primer indici potencial d'un comportament gregari fou el descobriment l'any 1878 de trenta-un Iguanodon que es pensà que havien mort junts a Bernissart, Bèlgica, després de caure en una dolina profunda i inundada, on s'havien ofegat.[53] Altres restes de morts en massa han estat descobertes des d'aleshores. Aquestes troballes, juntament amb moltes pistes de petges, suggereixen que el comportament gregari era habitual en moltes espècies de dinosaure. Pistes de petges de centenars o potser milers d'herbívors indiquen que els hadrosàurids es podrien haver mogut en grans ramats, com el bisó americà o la gasela saltadora. Les pistes de petges dels sauròpodes revelen que aquests animals viatjaven en grups composts de diverses espècies diferents, com a mínim a Oxford, Anglaterra,[54] tot i que no hi ha proves d'estructures de ramat específiques.[55] Els dinosaures podrien haver-se agrupat en ramats per defensar-se, per migrar o per protegir les cries. La interpretació dels dinosaures com a animals gregaris també s'ha estès als teròpodes carnívors, descrivint-los com a caçadors en grup que treballaven junts per abatre preses grans.[56][57] Tanmateix, aquest estil de vida no és habitual entre els parents moderns dels dinosaures (cocodrils i altres rèptils, juntament amb els ocells – l'aligot de Harris n'és una excepció ben documentada), i les proves tafonòmiques que semblen indicar un comportament de caça en grup entre teròpodes, com ara Deinonychus o l'al·losaure, també es poden interpretar com al resultat de disputes fatals entre animals que menjaven, com s'observa en molts depredadors diàpsids moderns.[58]

El descobriment de Jack Horner el 1978 d'una sèrie de nius de Maiasaura ("bona mare dinosaure") a Montana revelà que els ornitòpodes continuaven cuidant de les cries durant un període llarg després del naixement.[59] També hi ha proves que altres dinosaures del Cretaci, com els sauròpodes titanosaures de la Patagònia (descoberts el 1997) també aniuaven en grans grups.[60] L'oviraptòrid mongol Citipati fou descobert el 1993 en una posició similar a la de les gallines quan coven, cosa que podria significar que estava cobert d'una capa aïllant de plomes que mantenia la calor dels ous.[61] Altres descobriments també apunten que els pares cuidaven de les cries. Per exemple, s'han trobat les restes fòssils d'un grup de psitacosaures, consistent en un adult i trenta-quatre cries; en aquest cas, el gran nombre de cries es pot deure a un costum de fer nius comuns.[62] A més, es descobrí un embrió de dinosaure (pertanyent al prosauròpode Massospondylus) sense dents, indicant que era necessari un cert grau de cura pels pares per alimentar els dinosaures joves.[63] Les pistes de petges també han confirmat aquest comportament entre ornitòpodes de l'illa de Skye, al nord-oest d'Escòcia.[64] S'han trobat nius i ous de la majoria de grups principals de dinosaures, i sembla probable que els dinosaures es comuniquessin amb les cries, de manera similar als cocodrils i ocells actuals.

Les crestes d'alguns dinosaures, com ara els marginocèfals, teròpodes i lambeosaurins, podrien haver estat massa febles per ser utilitzades com una forma de defensa activa, de manera que probablement se les utilitzava per al festeig o per espantar els rivals, tot i que no se sap gaire sobre l'aparellament dels dinosaures o el seu territorialisme. Ferides de mossegades al cap suggereixen que, com a mínim, els teròpodes tenien confrontacions agressives actives.[65] La naturalesa de la comunicació dels dinosaures també roman enigmàtica, i és un àmbit actiu d'investigació. Per exemple, estudis recents suggereixen que les crestes buides dels lambeosaurins podrien haver funcionat com a cambres de ressonància, utilitzades per emetre una gran varietat de vocalitzacions.[66][67]
Un dels fòssils de dinosaure més valuosos per l'estudi del comportament d'aquests animals fou descobert al desert de Gobi el 1971. Consistia en un Velociraptor que atacava un Protoceratops,[68] oferint proves que, en efecte, els dinosaures s'atacaven els uns als altres.[69] Una altra prova que els dinosaures atacaven altres dinosaures vius és la cua parcialment curada d'un edmontosaure, un dinosaure hadrosàurid; la cua està danyada d'una manera tal que mostra que l'animal fou mossegat per un tiranosaure però sobrevisqué.[69] El canibalisme entre algunes espècies de dinosaure fou confirmat per marques de dents descobertes a Madagascar el 2003, associades al teròpode Majungasaurus.[70]
Basant-se en proves fòssils actuals de dinosaures com ara Oryctodromeus, sembla que algunes espècies herbívores tenien un estil de vida parcialment subterrani,[71] i algunes espècies semblants als ocells podrien haver estat arbòries, particularment dromeosàurids primitius com ara Microraptor[72] i els enigmàtics escansoriopterígids.[73] Tanmateix, sembla que molts dinosaures tenien un mode de locomoció terrestre. Una bona comprensió de com els dinosaures es movien a terra és clau per fer models del seu comportament; la ciència de la biomecànica, en particular, ha ofert indicis claus en aquest àmbit. Per exemple, estudis de la força exercida pels músculs i per la gravetat sobre l'estructura esqueletal dels dinosaures han servit per investigar a quina velocitat podien córrer,[74] si els diplodòcids podien crear bangs sònics espetegant la cua de manera semblant a un fuet,[75] i si els sauròpodes podien flotar.[76]
Fisiologia


Des de la dècada del 1960 hi ha hagut un intens debat sobre la regulació de la temperatura en els dinosaures. Originalment, els científics no es posaven gens d'acord sobre si els dinosaures eren capaços de regular la seva temperatura. Més recentment, l'homeotèrmia ha esdevingut la visió que gaudeix del major consens i el debat s'ha concentrat en els mecanismes termoreguladors.
Després del descobriment dels dinosaures, al principi els paleontòlegs sostingueren que es tractava de criatures ectoterms; «llangardaixos terribles», com diu el seu nom. Aquesta suposada ectotèrmia implicava que els dinosaures eren organismes relativament lents i inerts, de manera similar als rèptils moderns, que necessiten fonts externes de calor per regular la temperatura corporal. L'ectotèrmia dels dinosaures fou una visió prevalent, fins que Robert T. Bakker, un dels primers proponents de l'endotèrmia dels dinosaures, publicà un document influent sobre aquest tema el 1968.
Les proves modernes indiquen que els dinosaures prosperaren en climes freds, i que, com a mínim, algunes espècies de dinosaure regulaven la temperatura corporal per mitjà de processos biològics interns (potser amb l'ajut de la seva gran mida). Algunes proves d'endotèrmia en els dinosaures inclouen el descobriment de dinosaures polars a Austràlia i l'Antàrtida (on haurien experimentat un fred i fosc hivern de sis mesos), el descobriment de dinosaures les plomes dels quals haurien pogut funcionar com un aïllament termoregulador, i anàlisis d'estructures dels vasos sanguinis típiques d'animals endoterms dins els ossos de dinosaure. Les estructures esqueletals suggereixen que els teròpodes i altres dinosaures tenien estils de vida actius més aviat típics d'un sistema cardiovascular endotèrmic, mentre que els sauròpodes presentaven menys característiques endotèrmiques. És, sens dubte, possible que alguns dinosaures fossin endotèrmics i altres no. Encara continua el debat científic sobre certs aspectes d'aquest tema.[77]
El debat es complica pel fet que l'endotèrmia pot aparèixer basada en més d'un mecanisme. La majoria d'estudis sobre l'endotèrmia dels dinosaures tendeixen a comparar-los als ocells o mamífers mitjans, que gasten energies per elevar la temperatura corporal per sobre de l'ambiental. Els ocells i mamífers petits també tenen un aïllament que redueix la pèrdua de temperatura, com ara greix, pèl o plomes. Tanmateix, els mamífers grans, com ara els elefants, tenen un problema diferent a causa de la seva relació àrea de superfície-volum relativament baixa (principi d'Haldane). Aquesta relació compara el volum d'un animal amb l'àrea de la seva pell; a mesura que un animal augmenta de mida, la seva àrea de superfície augmenta més lentament que el seu volum. Arribat un cert punt, la quantitat de calor irradiada a través de la pell cau per sota de la quantitat de calor produïda dins del cos, obligant els animals a utilitzar mètodes alternatius per evitar la hipertèrmia. En el cas dels elefants, els adults tenen poc pèl, tenen grans orelles amb les quals incrementen la seva àrea de superfície, i també tenen adaptacions en el comportament (com ara l'ús de la trompa per ruixar-se amb aigua o el costum de banyar-se en el fang). Aquests comportaments augmenten la refrigeració per evaporació.
Es creu que els dinosaures grossos haurien hagut d'enfrontar-se a problemes similars; la seva mida corporal indica que perdien temperatura relativament lentament respecte a l'aire que els envoltava, de manera que podrien haver estat homeoterms inercials, animals que romanen més càlids que el seu ambient purament gràcies a la seva enorme mida, i no gràcies a adaptacions especials com les de les aus o els mamífers. Tanmateix, fins a present aquesta teoria no ha ofert una explicació vàlida en relació a les nombroses espècies de dinosaure de la mida d'un gos o d'una cabra, o les cries de les espècies més grosses.
Escanejades per mitjà de tomografia computada de la cavitat toràcica d'un dinosaure revelaren el que semblava les restes d'un cor de quatre cambres, similar al dels ocells i mamífers actuals.[78] Aquesta idea és controvertida dins de la comunitat científica, havent estat criticada com a ciència anatòmica defectuosa[79] o simplement una il·lusió.[80] La qüestió de com es relaciona aquest descobriment amb el ritme metabòlic i l'anatomia interna dels dinosaures pot ser discutible, tanmateix, poc importa la identitat de l'objecte: tant els crocodilians moderns com els ocells, els parents vivents més propers dels dinosaures, tenen cors amb quatre cambres (tot i que els dels crocodilians estan modificats), de manera que els dinosaures probablement també en tenien.[81]
Teixits tous i ADN

Un altre dels millors exemples d'impressions de teixit tou en un dinosaure fòssil fou descobert a Petraroia (Itàlia). El descobriment fou anunciat el 1998, i descrivia un exemplar d'un celurosaure petit i molt jove, Scipionyx samniticus. El fòssil incloïa parts dels intestins, el còlon, el fetge, els músculs, i la tràquea d'aquest dinosaure immadur.[43]
A l'edició del març del 2005 de la revista Science, la paleontòloga Mary Higby Schweitzer i el seu equip anunciaren el descobriment de material flexible similar als teixits tous dins de l'os de la cama d'un tiranosaure, amb una edat de 68 milions d'anys i provinent de la formació de Hell Creek de Montana. Després de descobrir-lo, l'equip rehidratà el teixit.[44]
Després de tractar l'os fossilitzat durant diverses setmanes per eliminar el contingut mineral de la cavitat fossilitzada de la medul·la òssia (un procés anomenat desmineralització), Schweitzer trobà proves d'estructures intactes, com ara vasos sanguinis, matriu òssia, i teixits connectius (fibres òssies). Una examinació sota el microscopi revelà que el suposat teixit tou de dinosaure havia conservat estructures fines (microestructures) fins i tot al nivell cel·lular. La naturalesa i composició exactes d'aquest material i les implicacions del descobriment de Schweitzer encara no estan clares; encara s'està estudiant i interpretant aquest material.[44]
Una investigació publicada a PloS One el 2018 ha posat en dubte les afirmacions que el material descobert és teixit tou de tiranosaure. Thomas Kaye, de la Universitat de Washington, i els seus coautors argumenten que el que realment omplia l'os de tiranosaure era simplement un biofilm viscós creat per bacteris, que omplirien els buits anteriorment ocupats pels vasos sanguinis i les cèl·lules.[82] Els investigadors descobriren que el que havia estat anteriorment identificat com a cèl·lules sanguínies, a causa de la presència de ferro, eren en realitat framboides, esferes minerals microscòpiques portadores de ferro. Trobaren esferes similars en una varietat d'altres fòssils de diversos períodes, incloent-hi un ammonit. En l'ammonit, trobaren les esferes en un lloc en què el ferro que contenen no podria haver tingut cap relació amb la presència de sang.[83]
S'ha anunciat en dues ocasions diferents l'extració amb èxit d'ADN antic de fòssils de dinosaure, però després d'una inspecció posterior i una avaluació d'experts, cap d'aquestes extraccions no ha pogut ser confirmada.[84] Tanmateix, s'ha inferit un pèptid visual funcional d'un dinosaure teòric per mitjà de l'ús de mètodes analítics de reconstrucció filogenètica en seqüències d'espècies properes modernes com ara rèptils o ocells.[85] A més, s'han detectat diverses suposades proteïnes en fòssils de dinosaure,[86] incloent-hi l'hemoglobina.[87]
Encara que es pogués reconstruir l'ADN dels dinosaures, seria excessivament difícil clonar i "criar" dinosaures amb la tecnologia actual, car no existeixen espècies suficientment properes com per utilitzar-ne els zigots, ni un ambient adient per al desenvolupament embrionari.
Plomes i origen dels ocells
La possibilitat que els dinosaures fossin els avantpassats dels ocells fou suggerida per primer cop el 1868 per Thomas Henry Huxley.[88] Després del treball de Gerhard Heilmann a principis del segle xx, la teoria dels ocells com a descendents dels dinosaures fou abandonada en favor d'avantpassats tecodonts, sent-ne la prova clau la suposada manca de clavícula dels dinosaures.[89] Tanmateix, com ho demostraren descobriments posteriors, les clavícules (o una única fúrcula fusionada, derivada de dues clavícules separades) no estaven realment absents;[24] ja se n'havia trobat el 1924 en Oviraptor, però havien estat identificades erròniament com a episternes.[90] En la dècada del 1970, John Ostrom recuperà la teoria dels dinosaures com a avantpassats de les aus,[91] que guanyà impuls durant les dècades següents amb l'adveniment de l'anàlisi cladística,[92] i un gran augment dels descobriments de petits teròpodes i ocells primitius.[12] Han estat d'especial rellevància els fòssils de la formació de Yixian, on s'han trobat múltiples teròpodes i ocells primitius, sovint amb plomes d'algun tipus.[24] Els ocells comparteixen més d'un centenar de trets anatòmics distints amb els dinosaures teròpodes, actualment considerats, generalment, els seus parents prehistòrics més propers.[93]
El grup més proper a ells és el dels celurosaures maniraptors.[24] Una minoria de científics, notablement Alan Feduccia i Larry Martin, han suggerit altres camins evolutius, incloent-hi versions revisades de la proposta de Heilmann dels arcosaures basals,[94] o que els teròpodes maniraptors eren els avantpassats dels ocells, però que no eren dinosaures, sinó convergents amb ells.[95]
Plomes

L'arqueòpterix, el primer bon exemple de «dinosaure amb plomes», fou descobert el 1860. L'exemplar original fou trobat als llims de Solnhofen al sud d'Alemanya, un jaciment rar i amb fòssils en un estat de conservació gairebé perfecte. L'arqueòpterix és un fòssil de transició, amb característiques clarament intermèdies entre les dels rèptils moderns i els ocells. Descobert només dos anys després de la influent obra de Darwin L'origen de les espècies, aquesta troballa intensificà el debat naixent entre els proponents de la biologia evolutiva i el creacionisme. Aquest ocell primitiu s'assembla tant als dinosaures que, sense una impressió clara de plomes a les roques que l'envoltaven, com a mínim un exemplar fou confós amb Compsognathus.[96]
Des de la dècada del 1990, s'ha descobert una sèrie de dinosaures amb plomes addicionals, oferint una prova encara més clara de l'estreta relació existent entre els dinosaures i els ocells actuals. La majoria d'aquests exemplars foren extrets del jaciment de la formació de Yixian, Liaoning (al nord-est de la Xina), que era part d'un continent insular durant el Cretaci. Tot i que només s'han trobat plomes en alguns indrets, és possible que els dinosaures no aviaris de la resta del món també en tinguessin. La manca de proves fòssils extenses de dinosaures no aviaris amb plomes es pot deure al fet que característiques delicades com ara la pell i les plomes, no solen quedar conservades pel procés de fossilització, de manera que estan absents del registre fòssil. Fins al present, es coneixen protoplomes (estructures primes semblants a filaments) de dinosaures a la base dels celurosaures; compsognàtids com Sinosauropteryx o tiranosauroïdeus com Dilong,[97] però plomes completes només es coneixen en el subgrup de celurosaures dels maniraptors, que inclou els oviraptorosaures, troodòntids, dromeosàurids, i ocells.[24][98] La descripció dels dinosaures amb plomes no ha mancat de controvèrsia; els crítics més importants han estat possiblement Alan Feduccia i Theagarten Lingham-Soliar, que han suggerit que les protoplomes són el resultat de la descomposició de la fibra col·laginosa que es trobava a sota del sistema integumentari dels dinosaures,[99][100][101] i que els maniraptors amb plomes completes no eren realment dinosaures, sinó convergents amb ells.[95] [100] Tanmateix, les seves visions no han estat acceptades en gran part per altres investigadors, fins al punt que s'ha posat en dubte la naturalesa científica de les afirmacions de Feduccia.[102]
Esquelet
Com que les plomes són associades sovint amb els ocells, els dinosaures amb plomes són titllats sovint descrits com l'anella perduda entre els ocells i els dinosaures. Tanmateix, les múltiples característiques esqueletals que comparteixen els dos grups també són un indici important pels paleontòlegs. Les àrees de l'esquelet amb semblances importants són el coll, el pubis, el canell (carp semilunar), el braç i la cintura escapular, la fúrcula i la quilla. La comparació dels esquelets de dinosaure i dels ocells per mitjà de l'anàlisi cladística reforça la teoria de la relació entre ambdós grups.
Un estudi del 2020 de la Universitat McMaster (Canadà) descobrí que els dinosaures també podien patir càncer després d'analitzar el peroné d'un centrosaure. Els científics determinaren que havia patit osteosarcoma, un càncer dels ossos.[103]
Teixits tous
Els grans dinosaures carnívors posseïen un complex sistema de sacs d'aire semblants als que tenen els ocells moderns, segons una investigació dirigida per Patrick O'Connor de la Universitat d'Ohio. És probable que els pulmons dels dinosaures teròpodes carnívors bípedes amb potes (similars a les dels ocells) bombegessin aire en sacs buits del seu esquelet, com passa en els ocells. «Allò que en el passat es considerava formalment un tret únic dels ocells estava present en certa forma en els avantpassats dels ocells», digué O'Connor.[104] En un article científic publicat a la revista en xarxa Public Library of Science ONE, els científcs descrigueren Aerosteon riocoloradensis, l'esquelet del qual ha proporcionat les proves més clares fins ara d'un dinosaure amb un sistema de respiració similar al dels ocells. Una tomografia computada posà de manifest la presència de sacs d'aire dins la cavitat corporal de l'esquelet d'Aerosteon.[105][106]
Una altra prova que els ocells i els dinosaures tenen una relació propera és l'ús de pedres al pedrer. Els animals s'empassaven pedres per fer més fàcil la digestió i per descompondre l'aliment i les fibres dures un cop entraven en l'estómac. Quan se les troba en associació amb fòssils, aquestes pedres reben el nom de gastròlits.[107]
Biologia reproductiva
El descobriment de certs trets en un esquelet de tiranosaure oferí recentment més proves que els dinosaures i les aus evolucionaren a partir d'un avantpassat comú i, per primer cop, permeté als paleontòlegs determinar el sexe d'un dinosaure. Quan ponen ous, les aus femelles desenvolupen un tipus especial d'os als membres entre el dur os exterior i la medul·la Aquest os "medul·lar", que és ric en calci, serveix per fer closques d'ou. La presència de teixits ossis derivats de l'endosti a les cavitats medul·lars interiors de parts de la pota posterior de l'exemplar de tiranosaure suggerí que Tyrannosaurus rex utilitzava estratègies reproductives similars, i revelà que l'exemplar era femella.[108] Investigacions posteriors han trobat os medul·lar en el teròpode Allosaurus i l'ornitòpode Tenontosaurus. Com que el llinatge de dinosaures que inclou l'al·losaure i el tiranosaure divergí del llinatge que inclou el tenontosaure molt aviat en l'evolució dels dinosaures, això suggereix que els dinosaures com a grup produïen teixit medul·lar. S'ha trobat os medul·lar en exemplars de mida subadulta, cosa que suggereix que els dinosaures assolien la maduresa sexual bastant aviat per animals tan grans.[109]
Proves sobre el comportament
Un fòssil de troodòntid recentment descobert demostra que alguns dinosaures dormien com alguns ocells moderns, amb el cap amagat sota el braç.[110] Aquest comportament, que podria haver contribuït a mantenir calent el cap, també és típic dels ocells moderns.
Extinció
Els dinosaures no aviaris s'extingiren sobtadament fa aproximadament seixanta-cinc milions d'anys. També s'extingiren molts altres grups d'animals, incloent-hi els ammonits (mol·luscs semblants als nàutils), mosasàurids, plesiosaures, pterosaures, tortugues herbívores, cocodrils, la majoria d'ocells, i molts grups de mamífers.[10] Aquesta extinció massiva rep el nom d'extinció del Cretaci-Paleogen. La naturalesa de l'esdeveniment que la causà ha estat estudiat intensament des de la dècada del 1970; actualment, existeixen diverses teories relacionades amb el suport dels paleontòlegs. Tot i que el consens general és que un impacte meteòric fou la causa principal de l'extinció dels dinosaures, alguns científics citen altres possibles causes, o donen suport a la idea que una combinació de diversos factors fou responsable de la sobtada desaparició dels dinosaures del registre fòssil.
Al punt àlgid del Mesozoic, no hi havia casquets polars, i el nivell del mar era entre 100 i 250 metres més alt que en l'actualitat. La temperatura del planeta també era molt més uniforme, amb una diferència de només 25 °C entre les temperatures polars i les equatorials. De mitjana, les temperatures atmosfèriques també eren molt més càlides; els pols, per exemple, eren 50 °C més càlids que avui en dia.[111][112]
La composició atmosfèrica durant el Mesozoic també era molt diferent. Els nivells de diòxid de carboni eren fins a dotze vegades superiors als de l'actualitat, i l'oxigen representava entre el 32-25% de l'atmosfera, en comparació amb el 21% d'avui dia. Tanmateix, a finals del Cretaci, l'ambient estava canviant dramàticament. Es reduí l'activitat volcànica, fet que provocà una tendència de refredament a mesura que els nivells de diòxid de carboni atmosfèric queien. Els nivells d'oxigen a l'atmosfera també començaren a fluctuar i acabaren caient considerablement. Alguns científics creuen que el canvi climàtic, juntament amb els nivells més baixos d'oxigen, podria haver provocat directament la desaparició de moltes espècies. Si els dinosaures tenien sistemes respiratoris similars als típics dels ocells moderns, podria haver estat particularment difícil per a ells adaptar-se a un rendiment respiratori inferior, tenint en compte l'enorme quantitat d'oxigen que necessitaven els seus cossos.[10]
Impacte meteòric

La teoria de la col·lisió amb un asteroide, que fou popularitzada el 1980 per Walter Alvarez i els seus col·legues, relaciona l'extinció de finals del període Cretaci amb un impacte amb un bòlid fa aproximadament 65,5 milions d'anys. Alvarez i altres suggeriren que un augment sobtat dels nivells d'iridi, registrat arreu del món en els estrats rocosos d'aquest període, era una prova directa de l'impacte.[113] La majoria dels indicis suggereixen actualment que un bòlid d'entre cinc i quinze quilòmetres de diàmetre impactà amb la Terra a prop de la península de Yucatán, creant el cràter de Chicxulub, de 170 km de diàmetre, i desencadenant l'extinció massiva.[114] Els científics no saben amb certesa si els dinosaures estaven prosperant o en declivi abans de l'impacte. Alguns científics suggereixen que el meteorit causà una baixada de la temperatura atmosfèrica llarga i no natural, mentre que d'altres, en canvi, afirmen que hauria provocat una ona de calor fora de l'habitual.
Tot i que el registre fòssil per si sol no permet inferir la rapidesa de l'extinció, alguns models suggereixen que fou extremament ràpida. El consens entre els científics que donen suport a aquesta teoria és que l'impacte provocà extincions tant directament (per la calor de l'impacte meteòric) com indirectament (per un refredament global a causa del fet que la matèria ejectada del cràter d'impacte reflectia la radiació tèrmica del Sol).
Al setembre del 2007, investigadors estatunidencs encapçalats per William Bottke del Southwest Research Institute de Boulder, i científics txecs utilitzaren simulacions per ordinador per identificar la font més probable de l'impacte de Chicxulub. Calcularen que hi havia un 90% de probabilitats que un asteroide gegant anomenat Baptistina, d'un diàmetre aproximat de 160 km, i que orbita en el cinturó d'asteroides entre Mart i Júpiter, rebés l'impacte d'un asteroide sense nom més petit, d'uns 55 km de diàmetre, fa 160 milions d'anys. L'impacte esberlà Baptistina, creant un grup que encara existeix, conegut actualment com a família Baptistina. Els càlculs indiquen que alguns dels fragments foren enviats directament en òrbites que es creuaven amb la de la Terra, un dels quals fou el meteorit de deu quilòmetres de diàmetre que impactà amb la península del Yucatán fa 65 milions d'anys, creant el cràter de Chicxulub.[115]
Tot i ser similar a la teoria de l'impacte d'Alvarez (que implicava un únic asteroide o cometa), aquesta teoria suggereix que "els passos de l'estrella companya del Sol, Nèmesi, a través del núvol d'Oort desencadenarien pluges de cometes."[116] Un o diversos d'aquests objectes col·lidiren posteriorment amb la Terra aproximadament al mateix temps, causant l'extinció global. Com en el cas d'un únic asteroide, el resultat final d'aquest bombardeig de cometes hauria estat una caiguda sobtada de les temperatures globals, seguida d'un prolongat període fred.[116]
Trapps del Dècan
Abans de 2000, els arguments que els trapps del Dècan causaren l'extinció estaven sovint relacionats amb la teoria que l'extinció fou gradual, car es pensava que els trapps començaren fa uns 68 milions d'anys i en duraren uns 2. Tanmateix, hi ha proves que dos terços dels trapps del Dècan foren creades en un milió d'anys, fa aproximadament 65,5 milions d'anys, de manera que aquestes erupcions haurien provocat una extinció bastant ràpida, possiblement en un període de milers d'anys, però igualment un període més llarg que el que caldria esperar-se d'un únic impacte meteòric.[117][118]
Els trapps del Dècan podrien haver causat l'extinció a través de diversos mecanismes, incloent-hi l'alliberament de pols i d'aerosols sulfúrics a l'aire, que podrien haver blocat la llum solar, reduint la capacitat fotosintètica de les plantes. A més, el vulcanisme dels trapps del Dècan podria haver provocat emissions de diòxid de carboni, que haurien augmentat l'efecte d'hivernacle una vegada la pols i els aerosols haguessin desaparegut de l'atmosfera.[118] Abans de l'extinció massiva dels dinosaures, l'alliberament de gasos volcànics durant la formació dels trapps del Dècan "contribuí a un escalfament global aparentment massiu. Algunes dades indiquen un augment mitjà de la temperatura d'aproximadament 8 °C en l'últim mig milió d'anys abans de l'impacte [a Chicxulub]."[117][118]
En els anys en què la teoria dels trapps del Dècan estava relacionada amb una extinció més lenta, Luis Alvarez (que morí el 1988), responia que els paleontòlegs estaven sent enganyats per les poques dades de què disposaven. Tot i que la seva afirmació no fou ben rebuda al principi, estudis de camp intensius posteriors dels estrats fossilífers reforçaren el seu argument. Finalment, la majoria de paleontòlegs començaren a acceptar la idea que les extincions en massa de finals del Cretaci es degueren principalment, o, com a mínim en part, a un impacte massiu de la Terra contra un bòlid. Tanmateix, fins i tot Walter Alvarez ha admès que hi hagué altres grans canvis a la Terra fins i tot abans de l'impacte, com ara un descens del nivell del mar i grans erupcions volcàniques que crearen els trapps del Dècan a l'Índia, i que aquests canvis podrien haver contribuït a les extincions.[119]
Manca d'adaptació a les condicions canviants
Al Cretaci mitjà, les plantes amb flor esdevingueren una part important dels ecosistemes terrestres, que anteriorment havien estat dominats per gimnospermes com ara les coníferes.[120] Els copròlits (fems fossilitzats) de dinosaure indiquen que, mentre que alguns menjaven angiospermes, la majoria de dinosaures herbívors s'alimentaven principalment de gimnospermes. Anàlisis estadístiques de Lloyd et al. arribaren a la conclusió que, a diferència del que deien estudis anteriors, els dinosaures no es diversificaren gaire al Cretaci superior. Lloyd et al. suggeriren que el fracàs dels dinosaures a l'hora d'adaptar-se a ecosistemes canviants els condemnà a l'extinció.[33]
Possibles supervivents al Paleocè
A vegades es troben restes de dinosaures no aviaris per sobre el límit K-Pg. El 2002, els paleontòlegs Zielinski i Budahn informaren del descobriment d'un únic fòssil d'un os de la cama d'un hadrosaure a la conca del San Juan (Nou Mèxic), i el descrigueren com una prova de dinosaures del Paleocè. La formació en què es descobrí l'os fou datada de principis de l'època del Paleocè, fa aproximadament 64,5 milions d'anys. Si l'os no fou redipositat en aquell estrat per l'acció de l'erosió, seria una prova que algunes poblacions de dinosaures podrien haver sobreviscut, com a mínim, mig milió d'anys dins l'era Cenozoica.[121] Altres proves inclouen el descobriment de restes de dinosaures a la formació de Hell Creek fins a 1,3 metres per sobre (equivalents a 40.000 anys després) del límit K-Pg. Han arribat descobriments similars d'altres llocs del món, incloent-hi la Xina.[122] Tanmateix, molts científics refusen que hi hagués hagut dinosaures al Paleocè, argumentant que aquests ossos foren ressituats, és a dir, que l'erosió els tragué de la seva ubicació inicial i quedaren enterrats de nou en sediments molt posteriors,[123][124] o pensen que, encara que fos certa, la presència d'uns quants dinosaures al Paleocè inferior no canviaria gaire cosa a efectes pràctics.[123]
Història del descobriment
Fa mil·lennis que es coneixen els fòssils de dinosaures, tot i que la seva autèntica naturalesa no era reconeguda. Els xinesos, que anomenen els dinosaures konglong (恐龍, o "drac terrible"), els consideraven ossos de drac i els documentaren com a tals. Per exemple, Hua Yang Guo Zhi, un llibre escrit per Zhang Qu durant la dinastia Jin Occidental, documentava el descobriment d'ossos de drac a Wucheng, a la província de Sichuan.[125] Els poblatans del centre de la Xina han excavat ossos de dinosaures des de fa dècades, pensant que pertanyien a dracs, per fer-ne medecines tradicionals.[126] A Europa, generalment es creia que els fòssils de dinosaures eren les restes de gegants i altres criatures que moriren al diluvi universal.

El megalosaure fou el primer dinosaure a ser descrit formalment, el 1677, quan se'n recuperà una part d'un os d'una pedrera de calcària a Cornualla, a prop de Chipping Norton, Anglaterra. Aquest fragment ossi fou identificat correctament com l'extrem inferior del fèmur d'un animal més gran que qualsevol ésser vivent en temps moderns. El segon gènere de dinosaure en ser identificat, Iguanodon, fou descobert el 1822 per Mary Ann Mantell, l'esposa del geòleg anglès Gideon Mantell. Gideon Mantell reconegué les semblances entre els seus fòssils i els ossos de les iguanes modernes. Dos anys més tard, el Reverend William Buckland, catedràtic de geologia a la Universitat d'Oxford, desenterrà més ossos fossilitzats de megalosaure i esdevingué la primera persona a descriure dinosaures en una revista científica.[127]
Aquests "grans llangardaixos fòssils" es convertiren ràpidament en un tema de gran interès pels científics europeus i estatunidencs, i el 1842 el paleontòleg anglès Richard Owen encunyà el terme "dinosaure". Descobrí que les restes que s'havien trobat fins aleshores, Iguanodon, Megalosaurus i Hylaeosaurus, comparteixen una sèrie de característiques distintives, de manera que decidí agrupar-los en un grup taxonòmic propi. Amb el suport del Príncep Albert de Saxònia-Coburg Gotha, marit de la Reina Victòria, Owen establí el Natural History Museum a South Kensington, Londres, per exhibir-hi la col·lecció nacional de fòssils de dinosaures i altres articles de biologia i geologia.
El 1858 es descobrí el primer dinosaure americà, en pous de marga al petit poble de Haddonfield (Nova Jersey) (tot i que se n'havien trobat fòssils abans, no se n'havia reconegut la naturalesa). La criatura fou denominada Hadrosaurus foulkii. Es tractava d'un descobriment extremament important: l'hadrosaure era un dels primers esquelets de dinosaure gairebé complets que es trobaven i era clarament una criatura bípeda (el primer havia estat descobert el 1834 a Maidstone, Anglaterra). Era un descobriment revolucionari car, fins aleshores, la majoria de científics pensaven que els dinosaures caminaven a quatre potes, com altres rèptils. Els descobriments de Foulke desencadenaren una onada de dinosauremania als Estats Units.[128]


Aquesta obsessió pels dinosaures quedava il·lustrada per la gran rivalitat entre Edward Drinker Cope i Othniel Charles Marsh, que competien per ser els primers a descobrir nous dinosaures en el que rebé el nom de Guerra dels Ossos. La rivalitat començà probablement quan Marsh remarcà públicament que la reconstrucció de Cope d'un esquelet d'elasmosaure tenia un defecte: Cope havia posat inadvertidament el cap del plesiosaure al lloc on hauria d'haver estat l'extrem caudal de l'animal. Aquest combat entre els dos científics durà més de trenta anys, arribant a la seva fi el 1897 quan Cope morí després de gastar-se tots els diners que tenia en la caça de dinosaures. Marsh "guanyà" la guerra principalment perquè estava més ben finançat gràcies a una relació amb el US Geological Survey. Desafortunadament, molts exemplars valuosos de dinosaures foren danyats o destruïts a causa dels mètodes bastos dels dos paleontòlegs; per exemple, els seus excavadors utilitzaven sovint dinamita per desenterrar ossos (un mètode que els paleontòlegs moderns trobarien atroç). Malgrat els seus mètodes bastos, les contribucions de Cope i Marsh a la paleontologia foren enormes; Marsh descobrí 86 espècies noves de dinosaure i Cope en trobà 56, fent un total de 142 espècies noves. La col·lecció de Cope es troba actualment al Museu Americà d'Història Natural de Nova York, mentre que la de Marsh està exposada al Museu Peabody d'Història Natural de la Universitat Yale.[129]
Des del 1897, la cerca de fòssils de dinosaures s'ha estès a tots els continents, incloent-hi l'Antàrtida. El primer dinosaure antàrtic que fou descobert, l'ancilosàurids Antarctopelta oliveroi, fou trobat a l'illa de Ross el 1986, tot i que no fou fins al 1994 que una espècie antàrtica, el teròpode Cryolophosaurus ellioti, fou descrit i anomenat formalment en una revista científica.
Els llocs d'especial investigació dels dinosaures en l'actualitat inclouen el sud de Sud-amèrica (particularment l'Argentina) i la Xina. La Xina, en particular, ha donat molts exemplars excepcionals de dinosaures amb plomes a causa de la geologia única dels seus estrats fossilífers, així com un antic clima àrid especialment favorable a la fossilització.
Renaixença dels dinosaures
El camp de la investigació dels dinosaures ha gaudit d'un augment de l'activitat que començà a la dècada del 1970 i encara continua. Això fou desencadenat, en part, pel descobriment de John Ostrom de Deinonychus, un predador actiu que podria haver estat de "sang calenta" (endotèrmia), en contrast amb la idea, aleshores prevalent, dels dinosaures com a animals lents de "sang fred" (poiquilotèrmia). La paleontologia dels vertebrats esdevingué un camp de la ciència global. Els paleontòlegs han fet grans descobriments de dinosaure en regions anteriorment ignorades, incloent-hi l'Índia, Sud-amèrica, Madagascar, l'Antàrtida, i particularment a la Xina (els dinosaures amb plomes sorprenentment ben conservats de la Xina han consolidat encara més la relació entre els dinosaures i els seus suposats descendents vivents, els ocells moderns). L'extensa aplicació de la cladística, que analitza rigorosament les relacions entre els organismes biològics, s'ha revelat extremament útil a l'hora de classificar els dinosaures. Les anàlisis cladístiques, juntament amb altres tècniques modernes, ajuden a compensar el fet que el registre fòssil és sovint incomplet i fragmentari.
Aspectes culturals

Des del punt de vista dels humans, els dinosaures eren criatures d'aparença fantàstica i sovint d'una mida enorme. Així, doncs, han atret la imaginació del públic i han esdevingut una part duradora de la cultura humana. Només tres dècades després de les primeres descripcions científiques de restes de dinosaures, s'erigiren unes cèlebres escultures de dinosaures al Crystal Palace Park de Londres. Aquestes cultures agradaren tant al públic que se'n vengueren rèpliques de mida inferior, un dels primers exemples de marxandatge de productes derivats. Des de la del Crystal Palace, s'han fet exhibicions de dinosaures a parcs i museus d'arreu del món, responent a l'interès públic i reforçant-lo.[130] La popularitat dels dinosaures també ha tingut un efecte recíproc sobre el coneixement dels dinosaures des de fa temps. La competició entre els museus per l'atenció del públic portà directament a la Guerra dels Ossos lliurada entre Marsh i Cope, cadascun intentant tornar amb restes fòssils més espectaculars que l'altre, i la contribució resultant al coneixement dels dinosaures fou enorme.[131]
Els dinosaures ocupen un lloc important a la cultura moderna. La paraula dinosaure es fa servir coloquialment per referir-se a quelcom de molt vell o molt gran. L'interès del públic pels dinosaures portà a la seva entrada inevitable a la cultura popular. Començant amb una menció de passada del megalosaure al primer paràgraf de Bleak House de Charles Dickens el 1852,[132] els dinosaures han aparegut en una gran varietat d'obres de ficció. El llibre d'Arthur Conan Doyle El món perdut, la pel·lícula icònica del 1933 King Kong, la pel·lícula de 1954 Godzilla i les seves múltiples seqüeles, el best-seller de 1990 Parc Juràssic i la seva versió en pel·lícula, que fou durant un curt temps la pel·lícula més taquillera de la història, són només alguns exemples prominents de la llarga tradició dels dinosaures en la ficció. Autors de no-ficció, incloent-hi alguns paleontòlegs prominents, també han intentat treure profit de la popularitat dels dinosaures, especialment entre els infants, per educar els lectors sobre els dinosaures en particular i la ciència en general. Els dinosaures són omnipresents en la publicitat, i moltes companyies intenten utilitzar els dinosaures per vendre els seus propis productes o per titllar els seus rivals com a lents o obsolets.[133]
Els dinosaures també han aparegut en els dibuixos animats, especialment després de la dinosauromania provocada per la pel·lícula de George Lucas i Steven Spielberg A la recerca de la vall encantada. La pel·lícula, que segueix les aventures d'un grup de joves dinosaures per arribar a la «Gran Vall», tingué tant d'èxit que al setembre de 2008, vint anys després de l'original, ja n'havien sortit dotze seqüeles. Altres programes infantils sobre dinosaures foren Denver, l'últim dinosaure, o Barney i amics. Els dinosaures també han aparegut a Power Rangers i, evidentment, a Els Picapedra. Finalment, cal remarcar que la reducció dels costs associats als efectes especials ha permès la creació de documentals televisius sobre els dinosaures, com ara Caminant entre dinosaures o When Dinosaurs Roamed America, que reprodueixen l'aspecte dels dinosaures amb una fidelitat sense precedents.
Concepcions religioses
Diversos grups religiosos tenen punts de vista sobre els dinosaures que difereixen dels que tenen la immensa majoria de científics, generalment a causa dels conflictes amb les històries de la Creació dels seus escrits sagrats. Tanmateix, la majoria de la comunitat científica refusa aquestes interpretacions dels dinosaures inspirades per la religió.[134][135]
Com que és un terme modern derivat del grec, la Bíblia no utilitza el mot "dinosaure", però el mot hebreu tanniyn /tan·ˈnin/ ha estat interpretat com una referència als dinosaures per alguns cristians.[136] En les traduccions al català, tanniyn es pot traduir com a "monstre marí" o "serpent", però se sol traduir com a "drac". Aquestes criatures són mencionades gairebé trenta vegades a l'Antic Testament, i es diu que viuen tant a la terra com a l'aigua. En un altre moment, la Bíblia descriu una criatura enorme anomenada behemot (Job 40:15-24), que "serva la cua tesa, com un cedre"; el behemot és descrit com "primera entre les obres de Déu" i impossible de capturar (v. 24). Alguns estudiosos de la Bíblia identifiquen el behemot com un elefant, un hipopòtam o un toro, però com que aquests animals tenen cues molt primes, no se les pot comparar a la mida d'un cedre; els creacionistes identifiquen sovint el behemot amb els dinosaures sauròpodes. Altres creacionistes es refereixen al behemot com a braquiosaure, car la Bíblia diu a Job que "és l'obra mestra de Déu", afirmant que és l'animal més gran que Déu creà.[136] Tanmateix, alguns estudiosos creuen que la referència al cedre es refereix en realitat a les seves fulles amb forma d'agulla, que s'assemblen al pèl punxegut present a la cua dels elefants, rinoceronts i hipopòtams d'avui en dia.[137] Altres crítics afirmen que el mot «cua» és un eufemisme pel penis de l'animal, i que s'ha d'entendre com una descripció de la seva virilitat.[138][139]
El leviatan és una altra criatura mencionada a l'Antic Testament; la seva descripció li atribueix una varietat de característiques de dinosaure, de drac, i de serp marina. Alguns estudiosos identifiquen el leviatan de Job 41 amb el cocodril del Nil, o assenyalen que té set caps i que és purament mitològic. Com en el cas del behemot, els creacionistes han intentat en algunes ocasions relacionar el leviatan amb els dinosaures.[140]
Referències
- ↑ Langer et al., 2010, p. 55.
- ↑ Bakker, R. T., Galton, P., 1974. "Dinosaur monophyly and a new class of vertebrates". Nature 248:168-172.
- ↑ R. Amiot, X. Wang, Z. Zhou, X. Wang, E. Buffetaut, C. Lecuyer, Z. Ding, F. Fluteau, T. Hibino, N. Kusuhashi, J. Mo, V. Suteethorn, Y. Wang, X. Xu, F. Zhang. Oxygen isotopes of East Asian dinosaurs reveal exceptionally cold Early Cretaceous climates. Proceedings of the National Academy of Sciences, 2011; DOI: 10.1073/pnas.1011369108
- ↑ Glut, Donald F. Dinosaurs: The Encyclopedia. Jefferson, North Carolina: McFarland & Co, 1997, p. 40. ISBN 0-89950-917-7.
- ↑ Lambert, David; and the Diagram Group. The Dinosaur Data Book. Nova York: Avon Books, 1990, p. 288. ISBN 978-0-380-75896-8.
- ↑ Morales, Michael. «Nondinosaurian vertebrates of the Mesozoic». A: Farlow, James O.; Brett-Surman, Michael K. (editors). The Complete Dinosaur. Bloomington: Indiana University Press, 1997, p. 607-624. ISBN 0-253-33349-0.
- ↑ Hu Yaoming; Meng Jin; Wang Yuanqing; and Li Chuankui «Large Mesozoic mammals fed on dinosaurs». Nature, 433, 2005, pàg. 149-152. DOI: 10.1038/nature03102.
- ↑ Wang, S.C., and Dodson, P. «Estimating the Diversity of Dinosaurs». Proceedings of the National Academy of Sciences USA, 103, 37, 2006, pàg. 13601-13605. DOI: 10.1073/pnas.0606028103. PMID: 16954187.
- ↑ Russell, Dale A. «China and the lost worlds of the dinosaurian era». Historical Biology, 10, 1995, pàg. 3-12.
- ↑ 10,0 10,1 10,2 MacLeod, N, Rawson, PF, Forey, PL, Banner, FT, Boudagher-Fadel, MK, Bown, PR, Burnett, JA, Chambers, P, Culver, S, Evans, SE, Jeffery, C, Kaminski, MA, Lord, AR, Milner, AC, Milner, AR, Morris, N, Owen, E, Rosen, BR, Smith, AB, Taylor, PD, Urquhart, E & Young, JR «The Cretaceous–Tertiary biotic transition». Journal of the Geological Society, 154, 2, 1997, pàg. 265-292. DOI: 10.1144/gsjgs.154.2.0265.
- ↑ 11,0 11,1 11,2 11,3 11,4 Benton, Michael J. «Origin and relationships of Dinosauria». A: Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (editors). The Dinosauria. 2a edició. Berkeley: University of California Press, 2004, p. 7-19. ISBN 0-520-24209-2.
- ↑ 12,0 12,1 Holtz, Jr., T.R.. Classification and evolution of the dinosaur groups. In: Paul, G.S. (editor). The Scientific American Book of Dinosaurs.. St. Martin's Press, 2000, p. 140-168. ISBN 0-312-26226-4.
- ↑ Langer, M.C., Abdala, F., Richter, M., and Benton, M.J. «A sauropodomorph dinosaur from the Upper Triassic (Carnian) of southern Brazil». Comptes Rendus de l'Academie des Sciences, Paris: Sciences de la terre et des planètes, 329, 1999, pàg. 511-517.
- ↑ Nesbitt, Sterling J.; Irmis, Randall B.; and Parker, William G. «A critical re-evaluation of the Late Triassic dinosaur taxa of North America». Journal of Systematic Palaeontology, 5, 2, 2007, pàg. 209-243. DOI: 10.1017/S1477201907002040.
- ↑ Això no es descobrí abans del 1909: «Dr. Holland and the Sprawling Sauropods». Arxivat de l'original el 2011-06-12. [Consulta: 14 agost 2008]. Els arguments i la majoria de les imatges també foren presentats a Desmond, A.. Hot Blooded Dinosaurs. DoubleDay, 1976. ISBN 0385270631.
- ↑ 16,0 16,1 Benton, M.J.. Vertebrate Paleontology. Blackwell Publishers, 2004, p. xii-452. ISBN 0-632-05614-2.
- ↑ Cowen, Richard. «Dinosaurs». A: History of Life. 4a ed.. Blackwell Publishing, p. 151-175. ISBN 1405117567.
- ↑ 18,0 18,1 Kubo, T.; M.J. Benton «Evolution of hindlimb posture in archosaurs: limb stresses in extinct vertebrates». Palaeontology, 50, 6, 2007, pàg. 1519-1529. DOI: 10.1111/j.1475-4983.2007.00723.x.
- ↑ Owen, R. (1842). "Report on British Fossil Reptiles." Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
- ↑ «Liddell-Scott-Jones Lexicon of Classical Greek». [Consulta: 5- 8-2008].
- ↑ Farlow, J.O.; Brett-Surman, M.K.. Preface. A: Farlow, J.O., and Brett-Surman, M.K. (editors). The Complete Dinosaur.. Indiana University Press, 1997, p. ix-xi. ISBN 0-253-33349-0.
- ↑ Irmis, Randall B. «A reappraisal of the phylogeny of early dinosaurs». The Palaeontological Association Newsletter, 65, 2007, pàg. 92-94 [Consulta: 22 novembre 2007].
- ↑ Olshevsky, G. (2000). "An annotated checklist of dinosaur species by continent." Mesozoic Meanderings, 3: 1-157
- ↑ 24,0 24,1 24,2 24,3 24,4 24,5 24,6 Padian, K. (2004). Basal Avialae. In: Weishampel, D.B., Dodson, P., i Osmólska, H. (editors). The Dinosauria (2a edició). University of California Press: Berkeley, 210-231. ISBN 0-520-24209-2
- ↑ Seeley, H.G. «On the classification of the fossil animals commonly named Dinosauria». Proc R Soc London. Royal Society, 43, 1887, pàg. 165-171.
- ↑ Romer, A. S.. Osteology of the Reptiles. University of Chicago, 1956.
- ↑ Ostrom, J.H. «Còpia arxivada». American Association for the Advancement of Science. Thomas, R.D.K. and Olson, E.C. [Boulder, CO], 1980, pàg. 82-105. Arxivat de l'original el 2008-05-27 [Consulta: 14 agost 2008]. Arxivat 2008-05-27 a Wayback Machine.
- ↑ Bakker, R. T., and Galton, P. «Dinosaur monophyly and a new class of vertebrates». Nature, 248, 1974, pàg. 168-172. DOI: 10.1038/248168a0.
- ↑ Kump LR, Pavlov A & Arthur MA «Massive release of hydrogen sulfide to the surface ocean and atmosphere during intervals of oceanic anoxia». Geology, 33, 5, 2005, pàg. 397–400. DOI: 10.1130/G21295.1.
- ↑ Tanner LH, Lucas SG & Chapman MG «Assessing the record and causes of Late Triassic extinctions». Earth-Science Reviews, 65, 1-2, 2004, pàg. 103-139. DOI: 10.1016/S0012-8252(03)00082-5.
- ↑ 31,0 31,1 Sereno PC «The evolution of dinosaurs». Science, 284, 5423, 1999, pàg. 2137–2147. DOI: 10.1126/science.284.5423.2137. PMID: 10381873.
- ↑ Sereno, P.C.; Forster, C.A.; Rogers, R.R.; and Monetta, A.M. «Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria». Nature, 361, 1993, pàg. 64–66. DOI: 10.1038/361064a0.
- ↑ 33,0 33,1 Lloyd, G.T., Davis, K.E., Pisani, D., et al «Dinosaurs and the Cretaceous Terrestrial Revolution». Proceedings of the Royal Society: Biology, 22-07-2008. DOI: 10.1098/rspb.2008.0715 [Consulta: 28 juliol 2008]. L'URL enllaça a tot el contingut, i és totalment lliure.
- ↑ Paul, G.S.. Predatory Dinosaurs of the World. Nova York: Simon and Schuster, 1988, p. 248-250.
- ↑ Clark, J.M., Maryanska, T., & Barsbold, R. (2004). "Therizinosauroidea", a The Dinosauria, 2nd ed. 151-164.
- ↑ Norell, M.A., & Makovicky, P.J. (2004). "Dromaeosauridae", a The Dinosauria, 2nd ed. 196-210.
- ↑ Daley, J. «New Study Restructures the Dinosaur Family Tree» (en anglès). Smithsonian.com, 24-03-2017. [Consulta: 26 març 2017].
- ↑ 38,0 38,1 38,2 38,3 Holtz, Thomas R., Jr.; Chapman, Ralph E.; & Lamanna, Matthew C.. «Mesozoic biogeography of Dinosauria». A: Weishampel, David B.; Dodson, Peter i Osmólska, Halszka (editors). The Dinosauria. 2a edició. Berkeley: University of California Press, 2004, p. 627-642. ISBN 0-520-24209-2.
- ↑ 39,0 39,1 39,2 39,3 39,4 Fastovsky, David E.; & Smith, Joshua B.. «Dinosaur paleoecology». A: Weishampel, David B.; Dodson, Peter i Osmólska, Halszka (editors). The Dinosauria. 2a edició. Berkeley: University of California Press, 2004, p. 614-626. ISBN 0-520-24209-2.
- ↑ Sereno, P.C.; Wilson, J.A.; Witmer, L.M.; Whitlock, J.A.; Maga, A.; Ide, O.; and Rowe, T.A. «Structural extremes in a Cretaceous dinosaur». PLoS ONE, 2, 11, 2007, pàg. e1230. DOI: 10.1371/journal.pone.0001230.
- ↑ Prasad, V.; Strömberg, C.A.E.; Alimohammadian, H; and Sahni, A. «Dinosaur coprolites and the early evolution of grasses and grazers». Science, 310, 5751, 2005, pàg. 1170-1180. DOI: 10.1126/science.1118806. PMID: 16293759.
- ↑ Archibald, J. David; and Fastovsky, David E.. «Dinosaur Extinction». A: Weishampel, David B.; Dodson, Peter i Osmólska, Halszka (editors). The Dinosauria. 2a edició. Berkeley: University of California Press, 2004, p. 672–684. ISBN 0-520-24209-2.
- ↑ 43,0 43,1 Dal Sasso, C.; Signore, M «Exceptional soft-tissue preservation in a theropod dinosaur from Italy». Nature, 292, 6674, 1998, pàg. 383-387. DOI: 10.1038/32884.
- ↑ 44,0 44,1 44,2 Schweitzer, M.H., Wittmeyer, J.L. and Horner, J.R. «Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex». Science, 307, 5717, 2005, pàg. 1952-1955. DOI: 10.1126/science.1108397. PMID: 15790853.
- ↑ «Assessment and Update Status Report on the Blue Whale Balaenoptera musculus». Committee on the Status of Endangered Wildlife in Canada, 2002. Arxivat de l'original el 2004-05-21. [Consulta: 5 desembre 2007].
- ↑ 46,0 46,1 Carpenter, Kenneth. «Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus». A: Foster, John R.; Lucas, Spencer G. (editors). Paleontology and Geology of the Upper Jurassic Morrison Formation (pdf). Albuquerque: New Mexico Museum of Natural History and Science, 2006, p. 131-138 (New Mexico Museum of Natural History and Science Bulletin 36) [Consulta: 16 agost 2008]. Arxivat 2007-12-02 a Wayback Machine.
- ↑ Farlow, James A. «On the rareness of big, fierce animals: speculations about the body sizes, population densities, and geographic ranges of predatory mammals and large, carnivorous dinosaurs». A: Dodson, Peter; and Gingerich, Philip. Functional Morphology and Evolution, 1993, p. 167-199 (American Journal of Science, Special Volume 293-A).
- ↑ Peczkis, J. (1994). "Implications of body-mass estimates for dinosaurs". Journal of Vertebrate Paleontology 14(4): 520-33
- ↑ «Anatomy and evolution». National Museum of Natural History. [Consulta: 21 novembre 2007].
- ↑ Colbert, E.H. (1968). Men and Dinosaurs: The Search in Field and Laboratory. E. P. Dutton & Company:New York, vii + 283 pàg. ISBN 0-14-021288-4
- ↑ Lovelace, David M.; Hartman, Scott A.; and Wahl, William R. «Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny». Arquivos do Museu Nacional, 65, 4, 2007, pàg. 527–544.
- ↑ dal Sasso, C., Maganuco, S., Buffetaut, E., and Mendez, M.A. (2006). New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities. Journal of Vertebrate Paleontology 25(4):888–896.
- ↑ Yans J, Dejax J, Pons D, Dupuis C & Taquet P «Implications paléontologiques et géodynamiques de la datation palynologique des sédiments à faciès wealdien de Bernissart (bassin de Mons, Belgique)» (en francès). Comptes Rendus Palevol, 4, 1-2, 2005, pàg. 135-150. DOI: 10.1016/j.crpv.2004.12.003.
- ↑ Day, J.J.; and Upchurch, P. «Sauropod trackways, evolution, and behavior». Science, 296, 2002, pàg. 1659. DOI: 10.1126/science.1070167. PMID: 12040187.
- ↑ Wright, Joanna L. «Steps in understanding sauropod biology». A: Curry Rogers, Kristina A.; Wilson, Jeffrey A.. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press, 2005, p. 252-284. ISBN 0-520-24623-3.
- ↑ Lessem, Don; and Glut, Donald F.. «Allosaurus». A: The Dinosaur Society's Dinosaur Encyclopedia. Random House, 1993, p. 19–20. ISBN 0-679-41770-2.
- ↑ Maxwell, W. D.; Ostrom, J.H. «Taphonomy and paleobiological implications of Tenontosaurus-Deinonychus associations». Journal of Vertebrate Paleontology, 15, 4, 1995, pàg. 707-712.(resum Arxivat 2007-09-27 a Wayback Machine.)
- ↑ Roach, Brian T.; and Brinkman, Daniel L. «A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs». Bulletin of the Peabody Museum of Natural History, 48, 1, 2007, pàg. 103-138. DOI: 10.3374/0079-032X(2007)48[103:AROCPH]2.0.CO;2.
- ↑ Horner, J.R.; and Makela, R. «Nest of juveniles provides evidence of family-structure among dinosaurs». Nature, 282, 5736, 1979, pàg. 296–298. DOI: 10.1038/282296a0.
- ↑ Chiappe, Luis M.; Jackson, Frankie; Coria, Rodolfo A.; & Dingus, Lowell. «Nesting titanosaurs from Auca Mahuevo and adjacent sites». A: Curry Rogers, Kristina A.; Wilson, Jeffrey A.. The Sauropods: Evolution and Paleobiology. Berkeley: University of California Press, 2005, p. 285-302. ISBN 0-520-24623-3.
- ↑ Oviraptor nesting Oviraptor nests or Protoceratops?
- ↑ Meng Qingjin; Liu Jinyuan; Varricchio, David J.; Huang, Timothy; and Gao Chunling «Parental care in an ornithischian dinosaur». Nature, 431, 2004, pàg. 145-146. DOI: 10.1038/431145a.
- ↑ Reisz RR, Scott, D Sues, H-D, Evans, DC & Raath, MA «Embryos of an Early Jurassic prosauropod dinosaur and their evolutionary significance». Science, 309, 2005, pàg. 761-764. DOI: 10.1126/science.1114942. PMID: 16051793.
- ↑ Dinosaur family tracks Les petges indiquen instint maternal després de deixar el niu.
- ↑ Tanke, Darren H.; and Currie, Philip J. «Head-biting behavior in theropod dinosaurs: paleopathological evidence» (pdf). Gaia, 15, 1998, pàg. 167-184. Arxivat de l'original el 2008-02-27. ISSN: 0871-5424 [Consulta: 18 agost 2008]. Arxivat 2008-02-27 a Wayback Machine.
- ↑ Hopson, James A «The evolution of cranial display structures in hadrosaurian dinosaurs». Paleobiology, 1, 1, 1975, pàg. 21–43.
- ↑ Diegert, Carl F.; Williamson, Thomas E «A digital acoustic model of the lambeosaurine hadrosaur Parasaurolophus tubicen». Journal of Vertebrate Paleontology, 18, 3, Suppl., 1998, pàg. 38A.
- ↑ «The Fighting Dinosaurs». American Museum of Natural History. [Consulta: 5 desembre 2007].
- ↑ 69,0 69,1 Carpenter, K. «Evidence of predatory behavior by theropod dinosaurs». Gaia, 15, 1998, pàg. 135-144. Arxivat de l'original el 2007-11-17 [Consulta: 5 desembre 2007]. Arxivat 2007-11-17 a Wayback Machine.
- ↑ Rogers, Raymond R.; Krause, David W.; & Curry Rogers, Kristina. «Cannibalism in the Madagascan dinosaur Majungatholus atopus». Nature, 422, 6931, 2007, pàg. 515–518. DOI: 10.1038/nature01532.
- ↑ Varricchio DJ, Martin, AJ and Katsura, Y «First trace and body fossil evidence of a burrowing, denning dinosaur». Proceedings of the Royal Society B: Biological Sciences, 274, 1616, 2007, pàg. 1361-1368. DOI: 10.1098/rspb.2006.0443.
- ↑ Chatterjee, S.; and Templin, R.J. «Biplane wing planform and flight performance of the feathered dinosaur Microraptor gui» (pdf). Proceedings of the National Academy of Sciences, 104, 5, 2007, pàg. 1576-1580. Arxivat de l'original el 2008-10-13. DOI: 10.1073/pnas.0609975104 [Consulta: 19 agost 2008].
- ↑ Zhang, F.; Zhou, Z.; Xu, X.; and Wang, X. «A juvenile coelurosaurian theropod from China indicates arboreal habits». Naturwissenschaften, 89, 9, 2002, pàg. 394–398. DOI: 10.1007 /s00114-002-0353-8.
- ↑ Alexander RM «Dinosaur biomechanics». Proceedings of the Royal Society of Biological Sciences, 273, 1596, 2006, pàg. 1849-1855. DOI: 10.1098/rspb.2006.3532.
- ↑ Goriely A & McMillen T «Shape of a cracking whip». Physical Review Letters, 88, 24, 2002, pàg. 244301. DOI: 10.1103/PhysRevLett.88.244301.
- ↑ Henderson, D.M. «Effects of stomach stones on the buoyancy and equilibrium of a floating crocodilian: A computational analysis». Canadian Journal of Zoology, 81, 8, 2003, pàg. 1346-1357. DOI: 10.1139/z03-122.
- ↑ Parsons, K.M. (2001). Drawing Out Leviathan. Indiana University Press. 22–48. ISBN 0-253-33937-5
- ↑ Fisher, P. E., Russell, D. A., Stoskopf, M. K., Barrick, R. E., Hammer, M. & Kuzmitz, A. A. «Cardiovascular evidence for an intermediate or higher metabolic rate in an ornithischian dinosaur». Science, 288, 5465, 2000, pàg. 503–505. DOI: 10.1126/science.288.5465.503. PMID: 10775107.
- ↑ Hillenius, W. J. & Ruben, J. A. «The evolution of endothermy in terrestrial vertebrates: Who? when? why?». Physiological and Biochemical Zoology, 77, 6, 2004, pàg. 1019-1042. DOI: 10.1086/425185.
- ↑ Rowe T, McBride EF, & Sereno PC «Dinosaur with a Heart of Stone». Science, 291, 5505, 2001, pàg. 783. DOI: 10.1126/science.291.5505.783a. PMID: 11157158.
- ↑ Chinsamy, Anusuya; and Hillenius, Willem J. (2004). "Physiology of nonavian dinosaurs". The Dinosauria, 2nd. 643–659.
- ↑ MKaye, TG; Gaugler G, Sawlowicz Z «Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms.». PLoS ONE, 3, 7, Juliol 2008, pàg. e2808. doi:10.1371/journal.pone.0002808 [Consulta: 29 juliol 2008].
- ↑ New Research Challenges Notion That Dinosaur Soft Tissues Still Survive Newswise, Consultat el 29 de juliol del 2008.
- ↑ Wang, H., Yan, Z. and Jin, D. «Reanalysis of published DNA sequence amplified from Cretaceous dinosaur egg fossil». Molecular Biology and Evolution, 14, 1997, pàg. 589–591. Arxivat de l'original el 2008-08-28 [Consulta: 5 desembre 2007]. Arxivat 2008-08-28 a Wayback Machine.
- ↑ Chang BS, Jönsson K, Kazmi MA, Donoghue MJ, Sakmar TP «Recreating a Functional Ancestral Archosaur Visual Pigment». Molecular Biology and Evolution, 19, 9, 2002, pàg. 1483-1489. PMID: 12200476 [Consulta: 5-12<2007].
- ↑ Embery G, Milner AC, Waddington RJ, Hall RC, Langley MS, Milan AM «Identification of proteinaceous material in the bone of the dinosaur Iguanodon». Connect Tissue Res, 44, Suppl 1, 2003, pàg. 41-6. DOI: 10.1080/713713598. PMID: 12952172.
- ↑ Schweitzer MH, Marshall M, Carron K, Bohle DS, Busse SC, Arnold EV, Barnard D, Horner JR, Starkey JR «Heme compounds in dinosaur trabecular bone». Proc Natl Acad Sci U S A., 94, 12, 1997, pàg. 6291-6. DOI: 10.1073/pnas.94.12.6291. PMID: 9177210.
- ↑ Huxley, Thomas H. «On the animals which are most nearly intermediate between birds and reptiles». Annals of the Magazine of Natural History, 4, 2, 1868, pàg. 66-75.
- ↑ Heilmann, Gerhard. The Origin of Birds. Londres: Witherby, 1926, p. 208pp.
- ↑ Osborn, Henry Fairfield «Three new Theropoda, Protoceratops zone, central Mongolia» (pdf). American Museum Novitates, 144, 1924, pàg. 1-12. Arxivat de l'original el 2009-03-27 [Consulta: 20 agost 2008].
- ↑ Ostrom, John H. «The ancestry of birds». Nature, 242, 5393, 1973, pàg. 136. DOI: 10.1038/242136a0.
- ↑ Gauthier, Jacques. «Saurischian monophyly and the origin of birds». A: Padian, Kevin. (ed.). The Origin of Birds and the Evolution of Flight, 1986, p. 1-55 (Memoirs of the California Academy of Sciences, 8).
- ↑ Mayr, G., Pohl, B. and Peters, D.S. «A Well-Preserved Archaeopteryx Specimen with Theropod Features». Science, 310, 5753, 2005, pàg. 1483-1486. DOI: 10.1126/science.1120331. PMID: 16322455.
- ↑ Martin, Larry D «A basal archosaurian origin for birds». Acta Zoologica Sinica, 50, 6, 2006, pàg. 977–990.
- ↑ 95,0 95,1 Feduccia, A. «Birds are dinosaurs: simple answer to a complex problem». The Auk, 119, 2002, pàg. 1187-1201. DOI: 10.1642/0004-8038(2002)119[1187:BADSAT]2.0.CO;2.
- ↑ Wellnhofer, P «Ein neuer Exemplar von Archaeopteryx». Archaeopteryx, 6, 1988, pàg. 1–30.
- ↑ Xu X.; Norell, M.A.; Kuang X.; Wang X.; Zhao Q.; and Jia C. «Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids». Nature, 431, 7009, 2004, pàg. 680-684. DOI: 10.1038/nature02855. PMID: 15470426.
- ↑ Göhlich, U.B.; and Chiappe, L.M. «A new carnivorous dinosaur from the Late Jurassic Solnhofen archipelago». Nature, 440, 2006, pàg. 329–332. DOI: 10.1038/nature04579.
- ↑ Lingham-Soliar, T. «The dinosaurian origin of feathers: perspectives from dolphin (Cetacea) collagen fibers». Naturwissenschaften, 12, 2003, pàg. 563–567. DOI: 10.1007/s00114-003-0483-7. PMID: 14676953.
- ↑ 100,0 100,1 Feduccia, A.; Lingham-Soliar, T.; and Hinchliffe, J.R. «Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence». Journal of Morphology, 266, 2, 2005, pàg. 125-166. DOI: 10.1002/jmor.10382. PMID: 16217748.
- ↑ Lingham-Soliar, T.; Feduccia, A.; and Wang X. «A new Chinese specimen indicates that 'protofeathers' in the Early Cretaceous theropod dinosaur Sinosauropteryx are degraded collagen fibres». Proceedings of the Biological Sciences, 274, 1620, 2007, pàg. 1823–9. DOI: 10.1098/rspb.2007.0352. PMID: 17521978.
- ↑ Prum, Richard O. «Are Current Critiques Of The Theropod Origin Of Birds Science? Rebuttal To Feduccia 2002». The Auk, 120, 2, Abril 2003, pàg. 550–61. DOI: 10.1642/0004-8038(2003)120[0550:ACCOTT]2.0.CO;2.
- ↑ Ekhtiari, Seper; Chiba, Kentaro; Popovic, Snezana; Crowther, Rhianne; Wohl, Gregory «First case of osteosarcoma in a dinosaur: a multimodal diagnosis» (en anglès). The Lancet Oncology, 21, 8, 01-08-2020, pàg. 1021-1022. DOI: 10.1016/S1470-2045(20)30171-6. ISSN: 1470-2045. PMID: 32758461.
- ↑ O'Connor PM & Claessens LPAM «Basic avian pulmonary design and flow-through ventilation in non-avian theropod dinosaurs». Nature, 436, 2005, pàg. 253–256. DOI: 10.1038/nature03716.
- ↑ Sereno, P.C.; Martinez, R.N.; Wilson, J.A.; Varricchio, D.J.; Alcober, O.A. i Larsson, H.C.E. «Evidence for Avian Intrathoracic Air Sacs in a New Predatory Dinosaur from Argentina». PLoS ONE, 3, 9, setembre 2008, pàg. e3303. DOI: 10.1371/journal.pone.0003303 [Consulta: 27 octubre 2008].
- ↑ Meat-Eating Dinosaur from Argentina Had Bird-Like Breathing System Newswise, consultat el 29 setembre 2008.
- ↑ Wings O «A review of gastrolith function with implications for fossil vertebrates and a revised classification» (Scholar search). Palaeontologica Polonica, 52, 1, 2007, pàg. 1-16 [Consulta: 24 novembre 2007].[Enllaç no actiu]
- ↑ Schweitzer, M.H.; Wittmeyer, J.L.; and Horner, J.R. «Gender-specific reproductive tissue in ratites and Tyrannosaurus rex». Science, 308, 2005, pàg. 1456-1460. DOI: 10.1126/science.1112158. PMID: 15933198.
- ↑ Lee, Andrew H.; and Werning, Sarah «Sexual maturity in growing dinosaurs does not fit reptilian growth models». Proceedings of the National Academy of Sciences, 105, 2, 2008, pàg. 582–587. DOI: 10.1073/pnas.0708903105. PMID: 18195356.
- ↑ Xu, X. and Norell, M.A. «A new troodontid dinosaur from China with avian-like sleeping posture». Nature, 431, 2004, pàg. 838–841. DOI: 10.1038/nature02898.
- ↑ Miller KG, Kominz MA, Browning JV, Wright JD, Mountain GS, Katz ME, Sugarman PJ, Cramer BS, Christie-Blick N, Pekar SF «The Phanerozoic record of global sea-level change». Science, 310, 5752, 2005, pàg. 1293–8. DOI: 10.1126/science.1116412. PMID: 16311326.
- ↑ McArthura JM, Janssenb NMM, Rebouletc S, Lengd MJ, Thirlwalle MF & van de Shootbruggef B «Palaeotemperatures, polar ice-volume, and isotope stratigraphy (Mg/Ca, δ18O, δ13C, 87Sr/86Sr): The Early Cretaceous (Berriasian, Valanginian, Hauterivian)». Palaeogeography, Palaeoclimatology, Palaeoecology, 248, 3-4, 2007, pàg. 391–430. DOI: 10.1016/j.palaeo.2006.12.015.
- ↑ Alvarez, LW, Alvarez, W, Asaro, F, and Michel, HV «Extraterrestrial cause for the Cretaceous–Tertiary extinction». Science, 208, 4448, 1980, pàg. 1095-1108. DOI: 10.1126/science.208.4448.1095. PMID: 17783054.
- ↑ Hildebrand, Alan R.; Penfield, Glen T.; Kring, David A.; Pilkington, Mark; Zanoguera, Antonio Camargo; Jacobsen, Stein B.; Boynton, William V. «Chicxulub Crater; a possible Cretaceous/Tertiary boundary impact crater on the Yucatan Peninsula, Mexico». Geology, 19, 9, Setembre 1991, pàg. 867-871. DOI: 10.1130/0091-7613(1991)019<0867:CCAPCT>2.3.CO;2.
- ↑ P, Claeys; Goderis, S «Solar System: Lethal billiards». Nature, 449, 05-09-2007, pàg. 30-31. DOI: 10.1038/449030a.
- ↑ 116,0 116,1 Koeberl, C & MacLeodKG. Geological Society of America. Catastrophic Events and Mass Extinctions, 2002. ISBN 0-8137-2356-6.
- ↑ 117,0 117,1 Hofman, C, Féraud, G & Courtillot, V «40Ar/39Ar dating of mineral separates and whole rocks from the Western Ghats lava pile: further constraints on duration and age of the Deccan traps». Earth and Planetary Science Letters, 180, 2000, pàg. 13-27. DOI: 10.1016/S0012-821X(00)00159-X.
- ↑ 118,0 118,1 118,2 Duncan, RA & Pyle, DG «Rapid eruption of the Deccan flood basalts at the Cretaceous/Tertiary boundary». Nature, 333, 1988, pàg. 841-843. DOI: 10.1038/333841a0.
- ↑ Alvarez, W. T. rex and the Crater of Doom. Princeton University Press, 1997, p. 130-146. ISBN 978-0691016306.
- ↑ Graeme T. Lloyd, Katie E. Davis, Davide Pisani, James E. Tarver, Marcello Ruta, Manabu Sakamoto, David W.E. Hone, Rachel Jennings, Michael J. Benton «Dinosaurs and the Cretaceous Terrestrial Revolution» (en anglès). Proceedings of the Royal Society, vol. 275, n. 1650, novembre 2008, p. 2483-2490.
- ↑ Fassett, JE, Lucas, SG, Zielinski, RA, and Budahn, JR «Compelling new evidence for Paleocene dinosaurs in the Ojo Alamo Sandstone, San Juan Basin, New Mexico and Colorado, USA». Catastrophic events and mass extinctions, Lunar and Planetary Contribution, 1053, 2001, pàg. 45-46 [Consulta: 18 maig 2007].
- ↑ Sloan, R. E., Rigby, K,. Van Valen, L. M., Gabriel, Diane «Gradual dinosaur extinction and simultaneous ungulate radiation in the Hell Creek Formation». Science, 232, 4750, 1986, pàg. 629-633. DOI: 10.1126/science.232.4750.629. PMID: 17781415 [Consulta: 18 maig 2007].
- ↑ 123,0 123,1 Fastovsky, David E.; and Sheehan, Peter M. «Reply to comment on "The Extinction of the dinosaurs in North America"» (pdf). GSA Today, 15, 2005, pàg. 11. DOI: 10.1130/1052-5173(2005)015[11b:RTEOTD]2.0.CO;2.
- ↑ Sullivan, RM «No Paleocene dinosaurs in the San Juan Basin, New Mexico». Geological Society of America Abstracts with Programs, 35, 5, 2003, pàg. 15. Arxivat de l'original el 2011-04-08 [Consulta: 2 juliol 2007]. Arxivat 2011-04-08 a Wayback Machine.
- ↑ Dong Zhiming. Dinosaurian Faunas of China. China Ocean Press, Beijing, 1992. ISBN 3-540-52084-8.
- ↑ «Dinosaur bones 'used as medicine'». BBC News, 2007. [Consulta: 6 juliol 2007].
- ↑ Sarjeant, William A.S.. «The earliest discoveries». A: Farlow, James O.; and Brett-Surman, Michael K. (editors). The Complete Dinosaur. Bloomington: Indiana University Press, 1997, p. 3-11. ISBN 0-253-33349-0.
- ↑ Prieto-Marques, A.; Weishampel, D.B.; Horner, J.R. «The dinosaur Hadrosaurus foulkii, from the Campanian of the East Coast of North America, with a reevaluation of the genus» (en anglès). Acta Palaeontologica Polonica, 51, 1, 2006, pàg. 77-98.
- ↑ Holmes T. Fossil Feud: The Bone Wars of Cope and Marsh, Pioneers in Dinosaur Science. Silver Burdett Press, 1996. ISBN 978-0382391477.
- ↑ Torrens, H.S. «The dinosaurs and dinomania over 150 years». Modern Geology, 18, 2, 1993, pàg. 257-286.
- ↑ Breithaupt, Brent H.. First golden period in the USA." In: Currie, Philip J. & Padian, Kevin (Eds.). The Encyclopedia of Dinosaurs. University of California Press, 1997, p. 347-350. ISBN 978-0122268106.
- ↑ "London. Michaelmas term lately over, and the Lord Chancellor sitting in Lincoln's Inn Hall. Implacable November weather. As much mud in the streets, as if the waters had but newly retired from the face of the earth, and it would not be wonderful to meet a Megalosaurus, forty feet long or so, waddling like an elephantine lizard up Holborne Hill." From page 1 of Dickens, Charles J.H. (1852). Bleak House. London: Bradbury & Evans.
- ↑ Glut, DF, & Brett-Surman, MK. Dinosaurs and the media." In: Farlow, James O. & Brett-Surman, Michael K. (Eds.). The Complete Dinosaur. Indiana University Press, 1997, p. 675-697. ISBN 978-0253213136.
- ↑ Kitcher, Philip. Abusing Science: The Case Against Creationism. MIT Press, 1983, p. 213. ISBN 978-0-262-61037-7 [Consulta: 31 agost 2008]. Arxivat 2008-06-23 a Wayback Machine.
- ↑ Dawkins, Richard. The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe Without Design. W.W. Norton, 1996, p. 400. ISBN 978-0393315707.
- ↑ 136,0 136,1 «Dinosaurs and the Bible». Clarifying Christianity', 2005. [Consulta: 14 març 2007].
- ↑ Bright, Michael. Beasts of the Field: The Revealing Natural History of Animals in the Bible, 2006, p346. 1861058314.
- ↑ Pennock, Robert T.. Tower of Babel: the evidence against the new creationism. Cambridge, Mass: MIT Press, 2000. ISBN 0-262-66165-9.
- ↑ «Index to Creationist Claims». [Consulta: 13 setembre 2007].
- ↑ «Index to Creationist Claims». the TalkOrigins Archive. [Consulta: 14 març 2007].
Bibliografia
- Langer, M. C.; Ezcurra, M. D.; Bittencourt, J. S.; Novas, F. E. «The origin and early evolution of dinosaurs» (en anglès). Biological Reviews, 85, 2010, pàg. 55-110. DOI: 10.1111/j.1469-185X.2009.00094.x.
Vegeu també
Enllaços externs
- Caminant entre dinosaures (anglès)